Kv1.3 channels participate in the activation and proliferation of leukocytes. The KCNE4 regulatory subunit associates with the channel and functions as a negative regulator. KCNE4, via its transmembrane and C-terminal domains, interacts with Kv1.3, impairing forward plasma membrane trafficking, decreasing macroscopic currents, and accelerating slow C-type inactivation of the channel. A negatively charged 145D/E polymorphic variant of KCNE4 has been associated with immune system disorders, such as allergic rhinitis and childhood acute lymphoblastic leukemia. In this work, we investigated the functional effects of these KCNE4 variants on Kv1.3 activity. Both variants similarly impaired forward trafficking of the channel. However, we observed a variant-dependent decrease in Kv1.3 currents with minor kinetic effects. In addition, we explored the effects of different residues at this position and analyzed the importance of the central amino acid of the polymorphism within the anionic (D–D/E–E) triplet. We suggest that the size and charge of the central position of the cluster are crucial for controlling Kv1.3 currents.
Introduction
The potassium channel regulatory subunit KCNE family comprises five members of single-spanning transmembrane domain (TMD) (type I) proteins (McCrossan and Abbott, 2004) that associate with different voltage-gated potassium (Kv) channels (Hille, 2001). KCNE regulation of Kv channels increases their functional diversity, enabling their participation in controlling gating kinetics, conductance, inactivation kinetics, plasma membrane trafficking, voltage dependence, and drug sensitivity (McCrossan and Abbott, 2004; Kanda and Abbott, 2012). The association of specific KCNE regulatory subunits with specific Kv channels is mandatory to recapitulate native currents, such as the cardiac IKs current generated by the association between KCNE1 and Kv7.1 (Sanguinetti et al., 1996; Lundquist et al., 2005). Evidence suggests that mutations in some KCNE members are strongly related to several human channelopathies (Sesti and Goldstein, 1998).
The Kv1.3 channel is essential to leukocyte physiology. KCNE4 regulates Kv1.3 (Grunnet et al., 2003). KCNE4 is the largest member of the KCNE family and has the longest C-terminal (Ct) domain. Both proteins are expressed in tissues, such as the immune system (Grissmer et al., 1990; Cahalan and Chandy, 2009; Vallejo-Gracia et al., 2021), brain, and muscle (Veh et al., 1995; Teng et al., 2003; Xu et al., 2004).
The main effect of KCNE4 on Kv1.3 activity is a substantial reduction in the current of the channel (Grunnet et al., 2003; Sole et al., 2009; Grunnet et al., 2020; Vallejo-Gracia et al., 2021; Sastre et al., 2024b). This inhibition is linked to KCNE4-dependent impairment of Kv1.3 plasma membrane targeting (Sole et al., 2009). Only KCNE4, out of the five KCNE family members, is able to modulate Kv1.3 trafficking (Sole et al., 2009; Sastre et al., 2024b). In addition to current amplitude, KCNE4 also regulates the characteristic slow C-type inactivation of the channel (Attali et al., 1992). KCNE4 accelerates this inactivation in a stoichiometry-independent mechanism (Sole et al., 2020) via interactions with the TMD and Ct domains of the regulatory protein (Sastre et al., 2024b).
Kv1.3 is important in leukocyte physiology for controlling T lymphocyte activation (Nguyen et al., 1996), proliferation (Conforti et al., 2003), and cytokine production (Hu et al., 2007). In this scenario, altered Kv1.3 activity is linked to autoimmune diseases such as multiple sclerosis (Beeton et al., 2001), rheumatoid arthritis (Beeton et al., 2006), or psoriasis (Kundu-Raychaudhuri et al., 2014). Consequently, Kv1.3 is of great therapeutic interest in the treatment of these disorders (Navarro-Perez et al., 2024). On the other hand, the role of KCNE4 in the pathogenesis of the immune system has been less explored. Kv1.3-associated T-cell physiological events are affected by changes in KCNE4 abundance (Vallejo-Gracia et al., 2021). The physiological role of KCNE4 is just emerging, and to completely understand the function of this ancillary subunit, further studies are needed.
In this work, we focused on a specific human KCNE4 polymorphism. The amino acid variation, which is located in the intracellular Ct domain of the protein, is correlated with immune system alterations such as childhood acute lymphoblastic leukemia (ALL) (Trevino et al., 2009) and allergic rhinitis (Freidin et al., 2013). This single-nucleotide polymorphism (SNP) affects a distal 145 position and involves an aspartic acid (Asp) residue (wild type [WT]) and a glutamic acid (Glu) in the polymorphic variant. Although both amino acids are similar with negatively charged side chains, we detected a variant-dependent reduction in Kv1.3 current. We analyzed how this KCNE4 minimal missense mutation affects Kv1.3 behavior. The effect of this SNP on Kv1.3, which is crucial in the immune system physiology, should be evaluated further in the context of the human KCNE4-related pathologies.
Materials and methods
Constructs
Human KCNE4 (short isoform) in pXOOM and human Kv1.3 in pRcCMV were generous gifts from J. Barhanin (Université Côte d’Azur, Nice, France) and F. Bezanilla (University of Chicago, Chicago, IL, USA), respectively. KCNE4 was subcloned and inserted into pECFP-N1 (Clontech Laboratories). Kv1.3 was subcloned and inserted into pEYFP-C1 (Clontech Laboratories) and pcDNA3. pDsRed-tagged pleckstrin homology domain of Akt, which is used as a transfectable membrane marker, was a gift from F. Viana (Universidad Miguel Hernández, Elche, Spain). KCNE4 mutants were generated using an Agilent QuikChange II site–directed mutagenesis kit and confirmed by Sanger sequencing.
Cell culture
Human embryonic kidney 293 (HEK293) cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum, 10,000 U/ml penicillin, and 100 μg/ml streptomycin (Thermo Fisher Scientific).
For electrophysiological experiments, cells were seeded in 35-mm dishes and transfected with lipotransfectin (Attendbio Research) according to the supplier’s instructions with 150 ng (Kv1.3) and 450 ng (KCNE4) of DNA. In the immunoprecipitation studies, HEK293 cells in a 100-mm dish were transfected with 4 μg of each DNA. For confocal imaging, cells were seeded on coverslips coated with poly-D-lysine and transfected with 750 ng of each DNA. All the experiments were performed 24–36 h after transfection.
Protein extraction, coimmunoprecipitation, and western blotting
Cells were washed twice in cold phosphate-buffered saline (PBS) and scraped in lysis buffer (1% Triton X-100, 10% glycerol, 50 mM HEPES, and 150 mM NaCl, pH 7.2) supplemented with the following protease inhibitors: 1 μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml pepstatin, and 1 mM PMSF (Sigma-Aldrich). The homogenates were subsequently centrifuged at 15,200 × g for 10 min, after which the supernatants were collected. The protein concentration was determined using Bio-Rad Bradford Assay Kit (Bio-Rad).
Up to 2 mg of protein was precleared with 50 μl of Protein A Sepharose beads (GE Healthcare) in 500 μl of washing buffer (150 mM NaCl, 50 mM HEPES, and 1% Triton X-100, pH 7.4) supplemented with protease inhibitors at 4°C with gentle mixing for 1 h. The precleared samples were incubated in Micro Bio-Spin Chromatography Column (Bio-Rad) for 2 h at room temperature (RT) with gentle mixing. The columns had been previously incubated for 1 h at RT with 50 μl of Protein A Sepharose beads crosslinked to 2.5 μl of anti-GFP antibody (GenScript) with 2.6 mg of dimethyl pimelimidate (Santa Cruz Biotechnology) for 30 min at RT with gentle mixing and washed with Tris-buffered saline. The columns were washed four times before elution with 0.2 M glycine (pH 2.5).
The samples were loaded on SDS–PAGE gels for protein electrophoresis and transferred to PVDF membranes (MilliporeSigma) and blocked in PBS with 0.05% Tween-20 and 5% dry milk for 1 h. The membranes were immunoblotted with antibodies against Kv1.3 (1/200; L23/27; NeuroMab) or GFP (1/200; 11814460001; Roche) overnight at 4°C or with β-actin (1/10,000; A5441; Sigma-Aldrich) at RT for 45 min. Blot images were obtained after the membranes were incubated with horseradish peroxidase anti-mouse secondary antibodies (1721011; Bio-Rad).
Confocal microscopy and image analysis
The seeded coverslips were washed twice with prewarmed (37°C) PBS without K+ and fixed for 10 min with prewarmed 4% paraformaldehyde (Sigma-Aldrich). Coverslips were mounted on microscope slides (Acefesa) with Mowiol-DABCO (Merck Millipore) and dried at RT for at least 3 days before imaging.
Images were obtained on a ZEISS LSM 880 laser scanning confocal spectral microscope (Carl Zeiss AG) using a 63× oil-immersion objective lens with a numerical aperture of 1.4. Image analysis was performed using ImageJ (National Institutes of Health). A pixel-by-pixel triple-colocalization analysis was performed using the plugin JACoP as previously described (Sastre et al., 2019), and Manders’ overlap coefficients (MOCs) were calculated.
Flow cytometry
The quantification of the fraction of protein at the plasma membrane was based on a previously published method (Bourdin et al., 2016). Nonpermeabilized HEK293 cells transfected with Kv1.3-hemagglutinin (HA) in the presence or absence of KCNE4-CFP constructs and resuspended in PBS were incubated with an anti-HA antibody (H6908; Sigma-Aldrich) conjugated with Cy5 at a 1:1,000 dilution at RT for 45 min. After washing, the presence of Kv1.3-HA at the plasma membrane was quantified as the Cy5 intensity in a BD FACSAria cell sorter. Cell viability was analyzed with propidium iodide staining. For data visualization, OMIQ (Dotmatics) software was used.
Electrophysiology
PatchMaster and FitMaster v2x90.5 (HEKA Elektronik GmbH) software packages were used for recording and analyzing the currents, respectively.
Statistical analysis
The results are expressed as individual values with the mean ± SE. Normality of the residuals was assessed using the Shapiro–Wilk test and the visual examination of the QQ plots. Homoskedasticity was addressed following the Brown–Forsythe test. Data from conditions following a normal distribution were analyzed with a one-way ANOVA and Tukey’s post hoc test. Welch’s ANOVA test with unpaired t test with Welch’s correction or Kruskal–Wallis and Dunn’s multiple comparisons test were used for experiments with significant P values for the normality and homoskedasticity tests. For coimmunoprecipitation (coIP) experiments, a repeated-measures one-way ANOVA was performed. For flow cytometry experiments, significance was calculated with a Brown–Forsythe ANOVA test. All statistical analyses were performed with GraphPad 8. Differences were considered significant when P < 0.05. A complete summary of statistical analysis is depicted in Table S1.
Online supplemental material
Fig. S1 shows confocal images of plasma membrane trafficking of the KCNE4 variants. Fig. S2 presents confocal microscopy images demonstrating that both KCNE4 (145D and 145E) variants similarly impair the plasma membrane targeting of Kv1.3. Fig. S3 analyzes the steady-state inactivation of Kv1.3 in the presence of KCNE4 variants. Table S1 summarizes the statistical analyses performed, including P values for the Shapiro–Wilk normality test, Brown–Forsythe test for homoskedasticity, and the statistical tests used to compare differences between conditions. Table S2 shows the voltage-dependent activation constant and τinactivation of Kv1.3 in the presence of KCNE4 variants. Table S3 presents the time constants for the activation and deactivation of Kv1.3 with the polymorphic KCNE4 variants. Table S4 shows the percentage of inactivation and τ of Kv1.3 current inactivation, as well as decay constants from a train of 15 depolarizing pulses.
Results
The 145D/E polymorphism of KCNE4 differentially affects the current density of Kv1.3
As previously reported (Grunnet et al., 2003; Sole et al., 2009), the presence of KCNE4 decreased Kv1.3 currents (Fig. 1 A) and accelerated slow C-type inactivation of the channel (Fig. 1 B). KCNE4 presents a nonsynonymous SNP in the Ct domain of the protein (rs12621643) resulting in a missense mutation from Asp (D) to Glu (E) at position 145 on hKCNE4. Although D represents the WT variant, E is the most prevalent residue at this position (Table 1). This SNP is localized in an intrinsically disordered region with a high density of negatively charged amino acids (Fig. 1, C and D). Different reports have correlated this polymorphism with the immune system such as childhood ALL (Trevino et al., 2009) and allergic rhinitis (Freidin et al., 2013), as well as cardiovascular disorders (Table 2). We first aimed to analyze how this KCNE4 minimal missense mutation affects Kv1.3 behavior. For that reason, we analyzed the effect of the KCNE4 145D/E polymorphism on Kv1.3 currents using the whole-cell patch clamp technique. Because the natural variants 145D and 145E preserve the negative charge, we also generated a neutral mutation to alanine (145A). Surprisingly, while KCNE4 145D decreased the Kv1.3 peak current density twofold, both 145E and 145A were responsible for the fourfold reduction in the current (Fig. 2, A–C). This difference in current density was not a result of changes in the voltage dependence of activation (Fig. 2 D and Table S2). Furthermore, the presence of either polymorphic KCNE4 variant did not affect the activation or deactivation kinetics of Kv1.3 (Table S3). KCNE4 variants accelerated slow C-type inactivation and decreased the remaining current at the end of a 250-ms-long pulse to a similar degree (Fig. 2 A and Table S4), as previously reported (Sastre et al., 2024b).
Kv1.3 plasma membrane trafficking is impaired by all KCNE4 variants
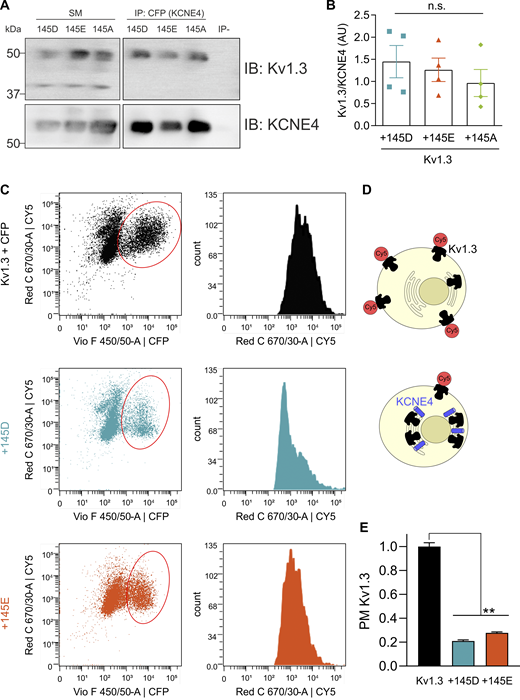
KCNE4 association impairs the forward plasma membrane trafficking of Kv1.3 (Sole et al., 2009; Sastre et al., 2024b). We analyzed whether the D/E polymorphism differentially altered the association between the channel and the regulatory subunit. CoIP (Fig. 3, A and B) and membrane localization (Fig. 3, C–E) studies were performed. In the coIP studies, we used a tagless Kv1.3 inserted in pcDNA3.1 and CFP-tagged KCNE4 (KCNE4-CFP). Kv1.3/KCNE4 coIP analysis revealed no differences among all KCNE4 mutants (Fig. 3, A and B). To assess plasma membrane localization of the channel, we performed flow cytometry experiments (Bourdin et al., 2016). HEK293 cells expressing Kv1.3 with an extracellular HA tag and different variants of KCNE4-CFP were used (Fig. 3 C). The cells were incubated with a primary antibody against HA (Kv1.3) and a secondary antibody conjugated to Cy5 (Fig. 3 D). The presence of both KCNE4 145D and 145E decreased the Cy5 signal in a similar manner, which correlated with lower levels of Kv1.3 at the plasma membrane (Fig. 3 E).
To further support these data, we performed confocal microscopy–based subcellular localization studies, in which we coexpressed Kv1.3-YFP, different KCNE4-CFP variants, and a membrane marker DsRed (Figs. S1 and S2). No differences in the intracellular retention were observed among the different KCNE4 mutants (Fig. S1). As previously described, pixel-by-pixel analysis of images revealed that Kv1.3, in the absence of KCNE4, mainly targeted the plasma membrane (Sole et al., 2009; Sastre et al., 2024b). However, the coexpression of all three KCNE4 variants similarly altered this distribution, retaining Kv1.3 intracellularly (Fig. S2 A). With a triple-colocalization assay (Sastre et al., 2019), MOC revealed that all KCNE4 variants decreased the plasma membrane targeting of Kv1.3 to a similar degree (Fig. S2 B). KCNE4 membrane expression did not differ between different mutants when it was expressed alone (Fig. S1) or in the presence of Kv1.3 (Fig. S2 C). Furthermore, no changes in Kv1.3/KCNE4 colocalization were observed for any KCNE4 variant (Fig. S2 D).
The polymorphism did not alter the KCNE4-dependent acceleration of slow inactivation of Kv1.3
KCNE4 not only promotes Kv1.3 retention in the endoplasmic reticulum (Sole et al., 2009; Sastre et al., 2024b) but also accelerates its characteristic slow C-type inactivation (Sole et al., 2020; Sastre et al., 2024b). Therefore, changes in the C-type inactivation between the variants could reduce the number of available channels, impacting their current density. Hence, we next examined the inactivation of the channel (Fig. 4 and Fig. S3) using long 5-s depolarizing pulses at +60 mV (Fig. 4, A and B). Currents were fitted to monoexponential equations to calculate the τinactivation (Fig. 4 C). All KCNE4 variants similarly accelerated the inactivation of Kv1.3, and no differences were detected (Table S4). KCNE4 also accelerates the use-dependent or cumulative inactivation of Kv1.3 (Sastre et al., 2024b). Applying a train of 15 depolarizing pulses (Fig. 4 D), the peak current of each pulse was fitted to a monoexponential equation (Fig. 4 E), and the decay constants were calculated (Fig. 4 F and Table S4). Similar to the slow C-type inactivation, all KCNE4s accelerated use-dependent inactivation to a similar degree. Therefore, modulation of channel inactivation is unlikely to be responsible for current density differences between KCNE4 variants.
Residue charge and position determine KCNE4 polymorphism effects
Although D/E substitutions represent minimal missense mutations, evidence suggests some pathological implications (Shapiro and Koshland, 1994; Brown et al., 2003; Parent et al., 2013; Debler et al., 2016). The KCNE4 145D/E SNP involves either an Asp or a Glu at the polymorphic position, two negatively charged residues whose side chains differ in length by one carbon. Because each variant decreases Kv1.3 currents in different magnitudes, we focused on the properties of the residue. Therefore, the polymorphic position was mutated to Lys (positively charged) and to Asn and Gln (polar uncharged residues). Kv1.3 current density in the presence of KCNE4 145K was within the range of KCNE4 (145D) (Fig. 5, A and B). However, the 145N and 145Q KCNE4 variants significantly decreased Kv1.3 currents.
Interestingly, the SNP is located within a highly anionic cluster of amino acids and is the central residue of a DDE motif (Fig. 5 C). The central Asp (D) is the polymorphic position and is surrounded by another Asp and a Glu. We investigated whether changes in these neighboring Asp and Glu positions also altered KCNE4 modulation of Kv1.3. Thus, we mutated the WT sequence (DDE) to DED and EDE (Fig. 5 C). Interestingly, we determined that solely the central amino acid of the triplet governed the current reduction (Fig. 5, D and E). Thus, unlike the WT (DDE) or alternative EDE, the positioning of an E in the center of the triplet (DED) reduced the Kv1.3 current by twofold over that of KCNE4 WT (Fig. 5 D).
Discussion
The KCNE families are important ion channel regulators, and mutations are involved in a plethora of diseases (Abbott and Goldstein, 2002). For example, for KCNE4, R20G is linked to atrial fibrillation (Gregers et al., 2017), whereas M58V is associated with thyrotoxic hypokalemic periodic paralysis (Silva et al., 2004). In this work, we investigated the effect of a polymorphism on Kv1.3 located in the distal Ct domain of KCNE4 (rs12621643, 145D/E) that is correlated with immune system alterations such as childhood ALL and allergic rhinitis (Trevino et al., 2009; Freidin et al., 2013). The potassium channel Kv1.3 is involved in the activation and proliferation of leukocytes, and KCNE4, being its ancillary subunit, is a strong modulator of the channel (Conforti et al., 2003; Hu et al., 2007). KCNE4 also regulates Kv7.1 (Ciampa et al., 2011), where the two KCNE4 variants were reported to have antagonistic effects on the channel (Ma et al., 2007); 145D increases Kv7.1 currents, whereas 145E triggers inhibition. However, the participation of the KCNE4 145D/E polymorphism in cardiac pathogenesis is the subject of debate (Hedley et al., 2011; Sinner et al., 2011).
In our findings, we observed differential regulation of the Kv1.3 current amplitude by the 145D/E KCNE4 polymorphism. Unlike on the Kv7.1 channel (Ma et al., 2007), both 145D and 145E produced a significant reduction in the current amplitude in Kv1.3, but the effect was twofold greater with KCNE4 145E. The KCNE4-dependent decrease in the Kv1.3 current has been linked mostly to impairment of forward plasma membrane trafficking of the channel (Sole et al., 2009; Sastre et al., 2024b). However, the presence of KCNE4 variants resulted in no apparent differences in the surface expression of Kv1.3. Because Kv1.3 channels possess a relatively large conductance, a subtle modulation of channel traffic can lead to larger-than-expected effects on the current density.
Although D/E is a minimal missense mutation, important pathogenic consequences have been reported, and differences in methylation claim alterations in protein structure (Shapiro and Koshland, 1994; Brown et al., 2003; Parent et al., 2013; Debler et al., 2016). In addition to KCNE4, natural D/E polymorphisms in other channel proteins have been described. In Kv11.1 (HERG), the E58D variant has a slower inactivation (Anderson et al., 2014), and E637D was identified as a risk variant for long QT syndrome (Amoros et al., 2011). We investigated some variations of the negatively charged residues within the 145-residue–containing triplet. By using the KCNE4 145A mutant, we determined that the presence of an Asp, rather than the absence of a Glu, was responsible for the changes. We also analyzed Kv1.3 currents in the presence of several KCNE4 mutants, such as 145K, 145Q, and 145N, at the polymorphic position. Only the currents in the presence of KCNE4 145K were comparable to those in the presence of WT 145D, whereas for KCNE4 145Q and 145N, the current density was significantly lower. These data suggest that the presence of a charged side chain, independent of its polarity, is necessary to limit how much KCNE4 reduces the current amplitude. The bulk and position of the charge could also impact how this residue interacts with others in KCNE4 or in the Kv1.3–KCNE4 complex. Because the polymorphism is located in a highly intrinsically disordered domain, predicting how this interaction could shape the overall structure of the complex is difficult (Punta et al., 2015; Bondos et al., 2022). In this structure, the polymorphic residue occupies the central position of a negatively charged D–D/E–E motif, and different Asp and Glu mutations within this triplet highlight the importance of the central position for determining the regulatory effect of KCNE4.
Considering the association between the variants and immune system diseases, we still need to examine how this interplay could affect leukocyte physiology. Interestingly, each variant is a risk factor for a different disease. KCNE4 145E is a risk factor for allergic rhinitis (Freidin et al., 2013), whereas 145D is associated with childhood ALL (Trevino et al., 2009). The pathogenesis of allergic rhinitis is mediated by several immune system cell types, including dendritic cells, B cells, and T cells (Zhang et al., 2022), and involves the production of IgE antibodies against inhaled allergens (Bousquet et al., 2020). Further research should explore how lower Kv1.3 activity, a consequence of the KCNE4 145E variant, could participate in this process and in which cell types this effect could be more relevant. On the other hand, in childhood ALL, KCNE4 145D could putatively produce higher Kv1.3 currents. This finding would be consistent with the pathophysiology of chronic lymphoblastic leukemia, in which the overexpression and overactivity of Kv1.3 contribute to disease (Leanza et al., 2013; Szabo et al., 2015), and pharmacological inhibition of Kv1.3 reduces tumor size (Severin et al., 2022). In summary, it is tempting to speculate that this minimal missense mutation, by controlling the Kv1.3 function, could trigger differential pathological outputs.
Data availability
Data are available in the article itself and its supplementary materials.
Acknowledgments
Christopher J. Lingle served as editor.
We thank Dr. Angela de Benito Bueno for her advice on cytometric methods, the CCiTUB Cytometry Facility, and Jaume Comas and Ricard Alvarez for their technical support.
The work was funded by the Ministerio de Ciencia, Innovación y Universidades (PID2023-148085OB-I00) and Agencia Estatal de Investigación (10.13039/501100011033), Spain and European Regional Development Fund, to A. Felipe. Open Access funding was provided by the Universitat de Barcelona.
Author contributions: Magalí Colomer-Molera: conceptualization, data curation, formal analysis, investigation, visualization, and writing—original draft, review, and editing. Daniel Sastre: conceptualization, formal analysis, visualization, and writing—review and editing. Antonio Felipe: conceptualization, data curation, funding acquisition, project administration, visualization, and writing—review and editing.
References
This work is part of a special issue on Emerging Research on Ion Channels in Health and Disease.