


Nerves are integral to the adenoid cystic carcinoma (ACC) microenvironment. The strong association of ACC with perineural invasion (PNI) is considered a hallmark of this disease. In human salivary ACC, we identify intratumoral, small-caliber, disorganized sympathetic nerves not observed in other salivary neoplasms. Norepinephrine or sympathetic ganglia explants enhance ACC proliferation in vitro. Two novel orthotopic ACC patient-derived xenograft (PDX) models recapitulate ACC morphology and demonstrate sympathetic innervation. Pharmacologic or surgical blockade of sympathetic nerves decreases ACC PDX growth. Bulk RNA sequencing of salivary ACC reveals correlations between noradrenergic nerve development signatures and worse patient survival. Metastatic ACC foci exhibit lower nerve signature gene expression levels than primary ACC. Sympathetic innervation in ACC is distinct from PNI and reflects tumor axonogenesis driven by noradrenergic neural development programs. These programs support ACC progression, are associated with poor prognosis, and may be inhibited as a therapeutic strategy.
Introduction
Adenoid cystic carcinomas (ACC) represent ∼10% of salivary gland cancers (Bradley, 2004). ACC are relatively slow-growing malignancies that can progress over years, frequently developing incurable late recurrences (Gondivkar et al., 2011). Regional nodal metastases are less frequent than distant metastases (Bradley, 2004). ACC has approximate 5-year and 10-year overall survival rates of 76% and 50%, respectively (Amit et al., 2015).
Perineural invasion (PNI), defined as cancer invasion in, around, and through nerves, is a hallmark characteristic of ACC (Amit et al., 2015; Rodriguez-Russo et al., 2021). Patients with ACC can present with pain, paresthesias, or paralysis, reflecting the spread of disease along nerves. Nerves microscopically associated with ACC are interpreted pathologically as PNI. Perineural tumor spread refers to the clinical and radiologic finding of cancer progression along or within large-caliber nerves (Bakst et al., 2019).
Tumor innervation is a contributor to tumor growth (Demir et al., 2021). Increased nerve density in breast (Pundavela et al., 2015), pancreatic (Ferdoushi et al., 2021), head and neck (Amit et al., 2020), and colorectal cancers (Albo et al., 2011) is correlated with poor prognosis. The autonomic nervous system enables prostate cancer initiation and progression (Magnon et al., 2013), while the parasympathetic nervous system promotes gastric cancer tumorigenesis and progression (Zhao et al., 2014). These findings opened a new field of cancer neuroscience (Demir et al., 2020, 2021; Amit et al., 2024; Hwang et al., 2025). Additional studies have also supported the contributions of tumor innervation to tumor growth. Targeting the nervous system reduces tumor growth in experimental models of breast (Kappos et al., 2018), gastric (Zhao et al., 2014), head and neck (Amit et al., 2020), pancreatic (Renz et al., 2018; Saloman et al., 2016), and prostate cancers (Magnon et al., 2013). Cancer-related stress and pain may also augment tumor progression through the sympathetic nervous system (Yonekura et al., 2022). Sympathetic nerves may have direct neurotransmitter effects on tumors or may stimulate angiogenesis (Zahalka et al., 2017). Therapy with β-blocker drugs has the potential to blunt sympathetic-driven tumor growth (Barron et al., 2011; Grytli et al., 2014; Renz et al., 2018).
The nearly ubiquitous identification of small nerves within ACC is pathologically labeled as PNI (Amit et al., 2015; Rodriguez-Russo et al., 2021). We hypothesized that this finding might alternately reflect tumor innervation. We postulate here that small nerves identified within ACC may represent new sympathetic nerve axonal growth and that this innervation impacts tumor progression. Established patient-derived xenograft (PDX) models of ACC allow for experimentation in the murine flank or zebrafish (Moskaluk et al., 2011) but have few nerves to study in the microenvironment. Here, we examine human ACC specimens and describe two new PDX models of ACC in the mouse submandibular salivary gland. We show that orthotopically implanted ACC PDX recapitulate histologic features of primary ACC and demonstrate sympathetic axonogenesis. The use of pharmacologic agents or surgical techniques blocks sympathetic innervation and inhibits tumor growth in ACC PDX. We demonstrate that sympathetic innervation of ACC supports tumor progression and suggest that the association of ACC with small sympathetic nerves reflects axonogenesis and tumor innervation, rather than PNI as conventionally interpreted. Deciphering the mechanisms through which neuronal signaling is co-opted to promote tumor growth may lead to new therapeutic approaches for patients with ACC.
Results
Human salivary ACC are innervated by large-caliber, intratumoral, sympathetic nerves
We performed immunofluorescence (IF) microscopy to identify and characterize the innervation of human salivary ACC specimens containing both tumor (T) and adjacent normal salivary gland tissue (N). We used tyrosine hydroxylase (TH) to identify sympathetic nerves, vasointestinal peptide (VIP) for parasympathetic nerves, and neurofilament heavy (NFH) or tubulin βIII (Tubb3) as a pan-neural marker (Fig. 1). Overlay of H&E and IF images show strong expression of intratumoral TH+ and VIP+ nerves surrounding cribriform areas. Intratumoral TH+ nerve staining is patchy and heterogeneous (Fig. 1 A).

Intratumoral sympathetic nerves in human salivary ACC. (A) ACC (T) intratumoral innervation and normal salivary gland (N). Immunostaining of sympathetic (TH), parasympathetic (VIP) nerves, and NFH for all nerves. Scale bar: low power, 20 μm; and high power, 50 μm. (B) Different innervation patterns between salivary ACC and normal acini. ACC: Disorganized intratumoral TH+ nerves located between the cribriform areas. Normal acini: Small-caliber, dot-like TH+ and VIP+ nerves encircle normal salivary acini. Tubb3 is a pan-neuronal marker. Scale bar: 50 μm. (C) TH+ nerve sprouts innervate ACC. ACC: Small-caliber TH+ nerves (arrowheads) arise from adjacent large-caliber TH+ nerves (dashed circles). Normal: No sprouts arise from large-caliber nerves (dashed circles). Tubb3 is a pan-neuronal marker representing all nerves on adjacent sections. Scale bar: low power, 200 μm; and high power, 50 μm. (D and E) IF and H&E staining of intratumoral TH+ nerves (D) and quantification (E) in salivary ACC (n = 10), normal salivary acini and hilum (n = 9), polymorphous adenocarcinoma (n = 3), mucoepidermoid carcinoma (n = 5), myoepithelial carcinoma (n = 4), acinic cell carcinoma (n = 8), and pleomorphic adenoma (n = 5). (A–D) Representative images from digitized whole-slide scans of composite 20× stitched images. Scale bar: 100 μm. P values were determined by one-way ANOVA and Fisher’s least significant difference (LSD) test. **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.
Intratumoral sympathetic nerves in human salivary ACC. (A) ACC (T) intratumoral innervation and normal salivary gland (N). Immunostaining of sympathetic (TH), parasympathetic (VIP) nerves, and NFH for all nerves. Scale bar: low power, 20 μm; and high power, 50 μm. (B) Different innervation patterns between salivary ACC and normal acini. ACC: Disorganized intratumoral TH+ nerves located between the cribriform areas. Normal acini: Small-caliber, dot-like TH+ and VIP+ nerves encircle normal salivary acini. Tubb3 is a pan-neuronal marker. Scale bar: 50 μm. (C) TH+ nerve sprouts innervate ACC. ACC: Small-caliber TH+ nerves (arrowheads) arise from adjacent large-caliber TH+ nerves (dashed circles). Normal: No sprouts arise from large-caliber nerves (dashed circles). Tubb3 is a pan-neuronal marker representing all nerves on adjacent sections. Scale bar: low power, 200 μm; and high power, 50 μm. (D and E) IF and H&E staining of intratumoral TH+ nerves (D) and quantification (E) in salivary ACC (n = 10), normal salivary acini and hilum (n = 9), polymorphous adenocarcinoma (n = 3), mucoepidermoid carcinoma (n = 5), myoepithelial carcinoma (n = 4), acinic cell carcinoma (n = 8), and pleomorphic adenoma (n = 5). (A–D) Representative images from digitized whole-slide scans of composite 20× stitched images. Scale bar: 100 μm. P values were determined by one-way ANOVA and Fisher’s least significant difference (LSD) test. **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.
Large-caliber TH+ nerves (>10 μm in diameter in the corresponding H&E sections) are more pronounced in ACC than in adjacent normal salivary gland tissue and demonstrate stronger staining than VIP+ nerves (Fig. 1 A) with a patchy, disorganized distribution pattern (Fig. 1, A and B). Small-caliber nerves were identified as single nerve fibers on IF imaging and were characterized with specific nerve markers.. Normal salivary glands have an organized pattern of small-caliber, dot-like TH+ and VIP+ nerves arranged around salivary acini (Fig. 1 B). Within ACC, large-caliber, intratumoral TH+ nerves are associated with a high density of small-caliber TH+ nerves in close proximity, suggesting that the small nerve fibers branch off the large-caliber nerves (Fig. 1 C, arrowheads). In contrast, IF microscopy of normal salivary glands shows large-caliber TH+ nerves only in the gland hila without adjacent small-calber TH+ nerves. (Fig. 1 C, white dashed circle). These asymmetric, disorganized nerve twigs and sprouts that arise near large-caliber TH+ nerves in ACC tumors are morphologically similar to new nerves and represent axonogenesis (Ayala et al., 2008; Baloh et al., 2022; Guillot et al., 2022; Madeo et al., 2018).
Large-caliber sympathetic innervation is absent in non-ACC salivary neoplasms
We compared TH+ sympathetic innervation in a panel of human salivary gland neoplasms. We quantified intratumoral, large-caliber TH+ nerves in ACC, polymorphous adenocarcinoma, mucoepidermoid carcinoma, myoepithelial carcinoma, acinic cell carcinoma, and pleomorphic adenoma (Fig. 1, D and E). Only ACC and polymorphous adenocarcinoma exhibited large-caliber intratumoral TH+ nerves, while other non-ACC salivary gland neoplasms rarely have intratumoral TH+ nerves. Large-caliber TH+ nerves are also identified in the hila of normal salivary glands (Fig. 1 D). The number of large-caliber TH+ nerves in ACC (0.634/mm2) is significantly higher in ACC compared with other types of salivary gland cancers (mucoepidermoid carcinoma: 0.0008/mm2, myoepithelial carcinoma: 0/mm2, acinic cell carcinoma: 0.0038/mm2, and pleomorphic adenoma: 0/mm2) (Fig. 1 E, P < 0.05 to 0.0001), but not polymorphous adenocarcinoma (0.2273/mm2, P not significant). TH+ nerve sprouts were also quantified, and only ACC, followed by polymorphous adenocarcinoma, demonstrated measurable nerve sprouts from this cohort of six salivary neoplasm types (Fig. S1).

Quantification of TH + nerve sprouts in human salivary gland neoplasms. Immunostaining of human salivary gland neoplasms (Fig. 1 E) was performed using TH antibody. The number of TH+ nerve sprouts per tumor area assessed was quantified. The density of TH+ nerve sprouts was highest in ACC (0.29 ± 0.085/mm2, n = 10), followed by polymorphous carcinoma (0.124 ± 0.029/mm2, n = 3). No or minimal TH+ nerve sprouts were identified in mucoepidermoid carcinoma (0.000 ± 0.000/mm2, n = 5), myoepithelial carcinoma (0.000 ± 0.000/mm2, n = 4), acinic cell carcinoma (0.006 ± 0.004/mm2, n = 8), or pleomorphic adenoma (0.000 ± 0.000/mm2, n = 5). P values were determined by one-way ANOVA and Fisher’s LSD test. **P < 0.01, ***P < 0.001, ns: not significant.
Quantification of TH + nerve sprouts in human salivary gland neoplasms. Immunostaining of human salivary gland neoplasms (Fig. 1 E) was performed using TH antibody. The number of TH+ nerve sprouts per tumor area assessed was quantified. The density of TH+ nerve sprouts was highest in ACC (0.29 ± 0.085/mm2, n = 10), followed by polymorphous carcinoma (0.124 ± 0.029/mm2, n = 3). No or minimal TH+ nerve sprouts were identified in mucoepidermoid carcinoma (0.000 ± 0.000/mm2, n = 5), myoepithelial carcinoma (0.000 ± 0.000/mm2, n = 4), acinic cell carcinoma (0.006 ± 0.004/mm2, n = 8), or pleomorphic adenoma (0.000 ± 0.000/mm2, n = 5). P values were determined by one-way ANOVA and Fisher’s LSD test. **P < 0.01, ***P < 0.001, ns: not significant.
Human salivary ACC exhibit β2-adrenergic receptor expression
We next investigated ACC cell expression of adrenergic receptors. Using ACC patient bulk RNA-sequencing (RNA-seq) datasets (Frerich et al., 2017; Linxweiler et al., 2020), we identified the β2-adrenergic receptor (β2R) as having the highest expression among adrenergic receptors β1, β2, and β3 (Fig. 2 A). Immunohistochemistry of β2R expression on a tissue microarray of human salivary ACC specimens reveals the majority of salivary ACC show moderate (9/28, 32.2%) to strong (13/28, 46.4%) expression of the β2R (Fig. 2 B). Only 6/28 (21.4%) of tumors show weak to negative expression.
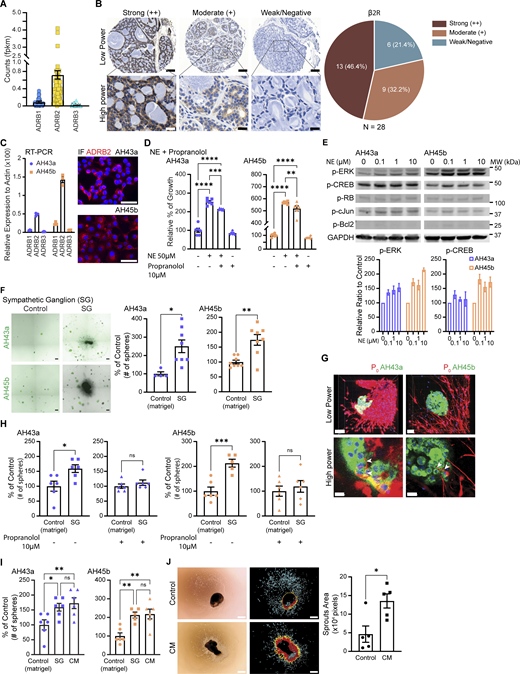
NE increases ACC cell proliferation in vitro. (A) Expression of β-adrenergic receptors from ACC RNA-seq datasets (n = 74). (B) Immunohistochemistry of β2R using a human salivary ACC tissue microarray (n = 28). Scale bar: low power, 100 μm; and high power, 20 μm. (C) RT-PCR of β-adrenergic receptors (n = 3 per group) and IF staining of β2R in AH43a and AH45b. Scale bar: 50 μm. (D) MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide] assay of AH43a and AH45b treated with NE, propranolol, or both (n = 6 per group). (E) Western blot of signaling pathways in AH43a and AH45b treated with NE at varying concentrations. Western blot quantification of pERK/GAPDH and pCREB/GAPDH compared with controls. (F) Tiled and stitched images of AH43a and AH45b spheroids cultured in Matrigel without (control) or with SG. Scale bar: 200 μm. Quantification of spheroid numbers for control and SG (n = 8 per group, 4 AH43a control). (G) Confocal images of P0+ TdTomato-lined SG neurites infiltrating into AH43a and AH45b spheroids (arrowheads show infiltrating neurites). Scale bar: low power, 100 μm; and high power, 20 μm. (H and I) Quantification of AH43a and AH45b spheroid numbers grown in Matrigel alone (control) or co-cultured with SG with propranolol or grown with CM from SG (n = 6 per group, 5 AH45b SG). (J) Left: Bright-field microscopy images of SG grown in regular media (control) or CM from AH43a after 7 days. Middle: Processed images showing the largest contiguous neurite area (red) emanating from the center (ganglion body). Right: Quantification of the neurite area (n = 5 per group). Data are representative of two independent experiments (D, F, and H–J). P values were determined by one-way ANOVA, Fisher’s LSD test (F and H), and Student’s t test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant (D and I). Source data are available for this figure: SourceData F2.
NE increases ACC cell proliferation in vitro. (A) Expression of β-adrenergic receptors from ACC RNA-seq datasets (n = 74). (B) Immunohistochemistry of β2R using a human salivary ACC tissue microarray (n = 28). Scale bar: low power, 100 μm; and high power, 20 μm. (C) RT-PCR of β-adrenergic receptors (n = 3 per group) and IF staining of β2R in AH43a and AH45b. Scale bar: 50 μm. (D) MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide] assay of AH43a and AH45b treated with NE, propranolol, or both (n = 6 per group). (E) Western blot of signaling pathways in AH43a and AH45b treated with NE at varying concentrations. Western blot quantification of pERK/GAPDH and pCREB/GAPDH compared with controls. (F) Tiled and stitched images of AH43a and AH45b spheroids cultured in Matrigel without (control) or with SG. Scale bar: 200 μm. Quantification of spheroid numbers for control and SG (n = 8 per group, 4 AH43a control). (G) Confocal images of P0+ TdTomato-lined SG neurites infiltrating into AH43a and AH45b spheroids (arrowheads show infiltrating neurites). Scale bar: low power, 100 μm; and high power, 20 μm. (H and I) Quantification of AH43a and AH45b spheroid numbers grown in Matrigel alone (control) or co-cultured with SG with propranolol or grown with CM from SG (n = 6 per group, 5 AH45b SG). (J) Left: Bright-field microscopy images of SG grown in regular media (control) or CM from AH43a after 7 days. Middle: Processed images showing the largest contiguous neurite area (red) emanating from the center (ganglion body). Right: Quantification of the neurite area (n = 5 per group). Data are representative of two independent experiments (D, F, and H–J). P values were determined by one-way ANOVA, Fisher’s LSD test (F and H), and Student’s t test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant (D and I). Source data are available for this figure: SourceData F2.
We assessed the expression of adrenergic receptors in two different human salivary ACC PDX, AH43a, and AH45b. Both cell types expressed adrenergic receptors, with the β2R showing the highest expression level (Fig. 2 C, left panel). IF microscopy showed strong membranous staining for the β2R (Fig. 2 C, right panel).
Adrenergic stimulation increases ACC proliferation in vitro
The addition of norepinephrine (NE) to AH43a and AH45b in vitro increased the proliferation of both cell types as compared with controls (Fig. 2 D, P < 0.0001). Pretreating the cells with the beta-blocker propranolol and then administering NE and propranolol in combination impairs the growth stimulation effect induced by NE alone (Fig. 2 D, P < 0.01 to 0.0001). AH43a and AH45b cells treated with increasing concentrations of NE showed increased p-ERK expression and smaller increases in p-CREB by western blot analysis (Fig. 2 E). Other potential markers of downstream β2R signaling pathways were unaffected, including p-c-Jun, p-RB, and p-Bcl2.
We assessed the impact of sympathetic neurotransmitter release on ACC spheroid growth in vitro. Because of NE’s short half-life, we used murine sympathetic ganglia (SG) co-cultured with ACC in 3D Matrigel culture (Fig. 2 F, left panel) to provide a continuous and sustained source of NE exposure over the 7-day period required for spheroid formation. We found a significantly increased number of tumor spheroids for AH43a and AH45b exposed to SG as compared with without exposure to SG (Fig. 2 F AH43a: control 13.625 ± 0.84, SG 23.75 ± 2.45 [174.3 ± 18% of control], P < 0.05; AH45b: control 2.25 ± 0.25, SG 5.625 ± 0.778 [250 ± 34% of control], P < 0.01).
We used confocal microscopy to examine interactions between ACC spheroids co-cultured in proximity to SG. Schwann cell-lined, small-caliber neurites extended from the SG and infiltrated into the ACC spheroids (Fig. 2 G). Under high power, the neurites appear to associate directly with individual ACC cancer cells in the spheroid (Fig. 2 G, lower panels, arrowheads). This suggests that ACC spheroids become innervated by neurites arising from the SG.
We co-cultured AH43a and AH45b tumor spheroids with or without SG in media with or without propranolol. In media without propranolol, both AH43a (159.07 ± 13.02% of control, P < 0.05) and AH45b (212.7 ± 15.9% of control, P < 0.001) co-cultured with SG showed a significant increase in spheroid numbers. In media with propranolol, propranolol abrogated SG’s proliferative effects in both AH43a (112 ± 9.12% of control, P not significant) and AH45b (119.24 ± 23.4% of control, P not significant) (Fig. 2 H). Propranolol likely exerts anti-adrenergic effects on both the ACC cells and the SG in this model (Du et al., 1993). We also found that conditioned media (CM) from SG significantly stimulated spheroid numbers as compared with controls (AH43a: 172.71 ± 19.92% of control, P < 0.01; AH45b: 218.16 ± 28.86% of control, P < 0.01) and no significant difference as compared with co-culture with SG (AH43a: 159.08 ± 14.26% of control; AH45b: 212.71 ± 17.78% of control) (Fig. 2 I). CM from cultured AH43a also stimulated SG neurite outgrowth as compared with regular media (Fig. 2 J, P < 0.05).
AH43a and AH45b PDX models recapitulate human salivary ACC
Both AH43a and AH45b PDX, generated from cultured single cancer cells, grow reliably in the salivary glands of mice and exhibit histologic characteristics of ACC, similar to the original tumor histology (Fig. 3 A), including areas of cribriform morphology and solid type pattern (Fig. 3 B), confirmed by a pathologist (N. Katabi). AH43a and AH45b cells both propagate in 2D as an epithelioid monolayer and in 3D Matrigel as tumor spheroids (Fig. 3 C).

Orthotopic ACC PDX recapitulates human ACC and derives sympathetic innervation. (A and B) H&E sections of patient specimens used to generate AH43a and AH45b and corresponding PDX. Scale bar: low power, 200 μm; and high power, 50 μm. (C) AH43a and AH45b cells grown in 2D and 3D cultures. Scale bar: 2D, 100 μm; and 3D, 50 μm. (D and E) Gross, ultrasound, and bioluminescence images and tumor volumes quantification of ACC PDX grown in the SMG, PG, or flank (n = 8). (F) TH+ innervation in ACC PDX (T) and adjacent normal salivary gland (N) of 20-μm sections. TH+ nerve fibers (arrowheads) sprout from large-caliber (dashed circles) TH+ nerves. Scale bar: low power, 100 μm; and high power, 50 μm. (G and H) Immunostaining of TH+ nerve sprouts (arrowheads) and large-caliber nerves (dashed circles) from ACC PDX in the mouse SMG and PG. Large-caliber nerves without nerve fiber sprouts are in the flank and lung. Quantification of intratumoral TH+ sprouts (AH43a, n = 5; AH45b, n = 8). Data are representative of two independent experiments (E and H). Scale bar: 50 μm. P values were determined by one-way ANOVA and Fisher’s LSD test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.
Orthotopic ACC PDX recapitulates human ACC and derives sympathetic innervation. (A and B) H&E sections of patient specimens used to generate AH43a and AH45b and corresponding PDX. Scale bar: low power, 200 μm; and high power, 50 μm. (C) AH43a and AH45b cells grown in 2D and 3D cultures. Scale bar: 2D, 100 μm; and 3D, 50 μm. (D and E) Gross, ultrasound, and bioluminescence images and tumor volumes quantification of ACC PDX grown in the SMG, PG, or flank (n = 8). (F) TH+ innervation in ACC PDX (T) and adjacent normal salivary gland (N) of 20-μm sections. TH+ nerve fibers (arrowheads) sprout from large-caliber (dashed circles) TH+ nerves. Scale bar: low power, 100 μm; and high power, 50 μm. (G and H) Immunostaining of TH+ nerve sprouts (arrowheads) and large-caliber nerves (dashed circles) from ACC PDX in the mouse SMG and PG. Large-caliber nerves without nerve fiber sprouts are in the flank and lung. Quantification of intratumoral TH+ sprouts (AH43a, n = 5; AH45b, n = 8). Data are representative of two independent experiments (E and H). Scale bar: 50 μm. P values were determined by one-way ANOVA and Fisher’s LSD test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.
AH43a or AH45b cells were injected into different microenvironments, including the submandibular gland (SMG), parotid gland (PG), and flank. Orthotopic tumors in either the SMG or the PG grew significantly larger than in the flank based on gross assessment (P < 0.001 to 0.0001), ultrasound assessment (P < 0.05 to 0.0001), and bioluminescence at week 3 (Fig. 3 D). We calculated the tumor volumes using small animal ultrasound and measured in vivo bioluminescence over 3 wk (Fig. 3 E). The SMG and PG tumors were significantly larger and had greater bioluminescence than the flank tumors. The SMG was overall the site most supportive for ACC progression, leading to its selection for tumor injections in subsequent experiments.
ACC PDX are innervated by large- and small-caliber, intratumoral, sympathetic nerves
We assessed the innervation of human salivary ACC PDX by staining for TH+ sympathetic nerves (Fig. 3 F) and identified tumor innervation patterns similar to the human ACC specimens (Fig. 1). IF microscopy revealed irregular, patchy areas of disorganized TH+ nerves at sites adjacent to the tumor-normal junction (Fig. 3 F, left). Similarly, at intratumoral locations, we observed multiple small-caliber, disorganized TH+ nerve fibers adjacent to large-caliber TH+ nerves. Small-caliber nerves (Fig. 3 F, middle, arrowheads) appeared to be sprouting off in multiple directions from large-caliber TH+ intratumoral nerves (Fig. 3 F, middle, dashed circles). This observation is consistent with similar findings in human salivary ACC surgical specimens (Fig. 1 C).
In contrast to ACC, TH+ nerves within the normal murine salivary gland form an organized, circular pattern surrounding normal acini (Fig. 3 F, right). This normal murine salivary sympathetic innervation pattern is consistent with normal human salivary tissue (Fig. 1, B and C). In the normal SMG, large-caliber TH+ nerve bundles can only be found in the hilar or interlobular areas (Fig. 3 F, right, dashed circles). Small-caliber TH+ nerve fibers do not appear to be emanating from adjacent large-caliber nerves.
Small-caliber axonogenesis is observed primarily in ACC PDX arising in SMG or PG sites (Fig. 3 G, arrowheads). Although large-caliber TH+ nerves are identified in ACC PDX in the flank or lung (Fig. 3 G, dashed circles), there are significantly greater numbers of small-caliber, disorganized TH+ nerve sprouts in the SMG as compared with flank or lung (Fig. 3 H, P < 0.05 for AH43a, P < 0.01 for AH45b). We also studied a non-ACC human salivary gland cancer, HTB-41. AH45b in the SMG showed significantly greater TH+ nerve sprouts than HTB-41 (Fig. S2 A, P < 0.01). There are no nerve sprouts seen arising from large-caliber intratumoral nerves in the HTB-41 tumors (Fig. S2 A, dashed circle), suggesting that axonogenesis is specific to ACC in salivary glands.

Reduced intratumoral nerve sprouting and no effect of sympathetic ablation in a non-ACC salivary cancer. (A) Large-caliber TH+ nerve (dashed circle) lacks any small caliber TH+ nerve sprouts in the non-ACC, mucoepidermoid salivary cancer, HTB-41, grown in the mouse SMG. Scale bar: 50 μm. Quantification of SMG intratumoral TH+ nerve sprouts for AH43a (n = 5), AH45b (n = 8), and HTB-41 (n = 8). AH43a and AH45b SMG TH+ data are derived from Fig. 3 H. (B) Volumes of HTB-41 tumors 3 wk after implantation in the mouse SMG, treated with either sham surgery (n = 9) or sympathectomy (n = 11). (C) RT-PCR of relative β2R expression in AH43a, AH45b, and HTB-41, using β-actin as endogenous control. Data represent one experiment (A–C). P values were determined by one-way ANOVA, Fisher’s LSD test (A), or Student’s t test (B). *P < 0.05; ns: not significant.
Reduced intratumoral nerve sprouting and no effect of sympathetic ablation in a non-ACC salivary cancer. (A) Large-caliber TH+ nerve (dashed circle) lacks any small caliber TH+ nerve sprouts in the non-ACC, mucoepidermoid salivary cancer, HTB-41, grown in the mouse SMG. Scale bar: 50 μm. Quantification of SMG intratumoral TH+ nerve sprouts for AH43a (n = 5), AH45b (n = 8), and HTB-41 (n = 8). AH43a and AH45b SMG TH+ data are derived from Fig. 3 H. (B) Volumes of HTB-41 tumors 3 wk after implantation in the mouse SMG, treated with either sham surgery (n = 9) or sympathectomy (n = 11). (C) RT-PCR of relative β2R expression in AH43a, AH45b, and HTB-41, using β-actin as endogenous control. Data represent one experiment (A–C). P values were determined by one-way ANOVA, Fisher’s LSD test (A), or Student’s t test (B). *P < 0.05; ns: not significant.
Pharmacologic or surgical ablation of murine salivary gland sympathetic innervation is effective
We used pharmacological and surgical methods to ablate sympathetic innervation in the murine salivary gland to assess the effect of TH+ nerves on cancer growth. We first assessed the efficiency of denervation. Chemical denervation with 6-OHDA leads to a dramatic reduction in the percentage of TH+ nerves as compared with all nerves (identified by Tubb3 staining) in normal murine salivary glands (Fig. S3 A, P < 0.0001). Surgical sympathectomy led to nearly complete ablation of TH+ nerves in normal salivary glands (Fig. S3 A, P < 0.0001). Both 6-OHDA and surgical sympathectomy showed highly effective TH+ nerve ablation at early (week 1) and late (week 3) time points. 6-OHDA–treated SMGs showed a mild recovery in TH+ nerve density but were still significantly lower than the control (Fig. S3, B and C, P < 0.05 to 0.01). Both treatments caused no significant changes in body weight, excised SMG weight, and cardiac output (Fig. S3 D). IF staining on these glands showed no significant difference in their CD31+ vessel density (Fig. S3 E), suggesting treatment-related hemodynamic or vascular changes are unlikely to account for observed tumor reduction. Pharmacologic inhibition of the β2R with propranolol did not alter the percentage of TH+ nerves in the normal salivary gland (Fig. S3 A). Sensory (TRPV1+) and parasympathetic (VIP+) nerves were not affected by surgical sympathectomy (Fig. S3 F).

Validation of sympathetic ablation of normal murine salivary glands showing no impact on vascularity or physiological parameters. (A) TH staining of normal murine submandibular salivary glands treated with propranolol (n = 6), 6-OHDA (n = 5), or surgical cervical sympathectomy (n = 7, control n = 15). Scale bar: 50 μm. Quantification of TH+ nerves in normal salivary glands as the percentage of TH+ nerves out of all Tubb3+ nerves in the above conditions. (B and C) TH staining of SMG treated with 6-OHDA (B) (week 1: n = 8 control and 4 6-OHDA; week 3: n = 6 control and 3 6-OHDA) and unilateral cervical sympathectomy (sympathetic cervical ganglionectomy [SCGx]) (C) (week 1: n = 5 per group, week 3: n = 3 per group) at early (week 1) and late (week 3) time points. Scale bar: 200 μm. Quantification of TH+ nerve density as TH+ area over selected area. (D) Body weight (left panel) (week 0–1: n = 6 control, 7 6-OHDA, and 8 SCGx; week 2–3: 3 per group), excised SMG weight (middle panel) (week 1: n = 3 control, 4 6-OHDA, and 5 SCGx; week 3: n = 3 per group) in mice treated with either 6-OHDA or SCGx at week 0, and cardiac output measured by LV ejection fraction at week 3 (right panel) (n = 3 per group). (E) IF of TH and CD31 of sections from each treatment at week 1 and week 3 (left panel). Quantification of CD31+ vessel density showed no significant difference for either treatment as compared with control at each time point (week 1: n = 8 control, 4 6-OHDA, and 5 SCGx; week 3: n = 6 control, 3 6-OHDA, and 3 SCGx). (F) (20 μm sections) TH co-localizes with Tubb3 in sham-treated salivary gland and has diminished expression in the sympathectomized gland. VIP and TRPV1 co-localize with TUBB3 and NFH and maintain expression in both sham and sympathectomized mouse salivary glands. Scale bar: 50 μm. Data are representative of two independent experiments (A–C and E) or one experiment (D). P values were determined by one-way ANOVA, Fisher’s LSD test (A, D, and E), or Student’s t test (B and C). *P < 0.05, **P < 0.01, ****P < 0.0001, ns: not significant.
Validation of sympathetic ablation of normal murine salivary glands showing no impact on vascularity or physiological parameters. (A) TH staining of normal murine submandibular salivary glands treated with propranolol (n = 6), 6-OHDA (n = 5), or surgical cervical sympathectomy (n = 7, control n = 15). Scale bar: 50 μm. Quantification of TH+ nerves in normal salivary glands as the percentage of TH+ nerves out of all Tubb3+ nerves in the above conditions. (B and C) TH staining of SMG treated with 6-OHDA (B) (week 1: n = 8 control and 4 6-OHDA; week 3: n = 6 control and 3 6-OHDA) and unilateral cervical sympathectomy (sympathetic cervical ganglionectomy [SCGx]) (C) (week 1: n = 5 per group, week 3: n = 3 per group) at early (week 1) and late (week 3) time points. Scale bar: 200 μm. Quantification of TH+ nerve density as TH+ area over selected area. (D) Body weight (left panel) (week 0–1: n = 6 control, 7 6-OHDA, and 8 SCGx; week 2–3: 3 per group), excised SMG weight (middle panel) (week 1: n = 3 control, 4 6-OHDA, and 5 SCGx; week 3: n = 3 per group) in mice treated with either 6-OHDA or SCGx at week 0, and cardiac output measured by LV ejection fraction at week 3 (right panel) (n = 3 per group). (E) IF of TH and CD31 of sections from each treatment at week 1 and week 3 (left panel). Quantification of CD31+ vessel density showed no significant difference for either treatment as compared with control at each time point (week 1: n = 8 control, 4 6-OHDA, and 5 SCGx; week 3: n = 6 control, 3 6-OHDA, and 3 SCGx). (F) (20 μm sections) TH co-localizes with Tubb3 in sham-treated salivary gland and has diminished expression in the sympathectomized gland. VIP and TRPV1 co-localize with TUBB3 and NFH and maintain expression in both sham and sympathectomized mouse salivary glands. Scale bar: 50 μm. Data are representative of two independent experiments (A–C and E) or one experiment (D). P values were determined by one-way ANOVA, Fisher’s LSD test (A, D, and E), or Student’s t test (B and C). *P < 0.05, **P < 0.01, ****P < 0.0001, ns: not significant.
Sympathetic ablation reduces ACC tumor volume
We inhibited the sympathetic nervous system in the murine salivary gland and assessed orthotopic ACC PDX tumor growth. Chemical ablation of sympathetic nerves with 6-OHDA (Fig. 4 A, P < 0.001), and surgical ablation of the cervical SG (Fig. 4 B, P < 0.001) resulted in significantly decreased tumor volumes for both AH43a and AH45b at 3 wk after tumor cell injection. In contrast, a non-ACC salivary cancer (HTB-41) did not show any tumor volume difference as compared with control (Fig. S2 B). Surgical sympathectomy exhibited the strongest suppressive effect on AH43a and AH45b tumor growth and appeared consistent with a greater degree of TH+ nerve ablation as compared with 6-OHDA ablation (Fig. S3). β2R blockade with propranolol also significantly reduced AH43a and AH45b PDX tumor volumes compared with control (Fig. 4 C, P < 0.001). We also created a CRISPR knockdown of the β2R (sgADRB2) in both AH43a and AH45b cells, confirmed by RT-PCR (Fig. S4 A). Isoproterenol stimulation showed that its proliferative effects were diminished in sgADRB2 as compared with nontargeted (NT) controls (Fig. S4 B). PDX sgADRB2 tumors from AH43a and AH45b were significantly smaller than their NT controls (Fig. 4 D, P < 0.05). These findings demonstrate that adrenergic stimulation supports ACC growth.

Sympathetic nerve ablation slows ACC growth in vivo. (A–D) AH43a and AH45b tumor volumes from mice treated with 6-OHDA, cervical sympathectomy, propranolol, or sgADRB2 knockdown and NT control at week 3. (E–G) NE concentrations in AH43a and AH45b tumors treated with 6-OHDA, cervical sympathectomy, and propranolol. (H–J) TH+ nerves in AH43a and AH45b tumors and quantification of the percentage TH+ nerves out of all Tubb3+ nerves following propranolol, 6-OHDA, cervical sympathectomy, and sgADRB2 and NT control tumors of AH43a and AH45b at week 3. Scale bar: 50 μm. (K–N) Immunostaining and quantification of CD31+ microvessel length per DAPI+ area in AH43a and AH45b tumors treated with propranolol, 6-OHDA, cervical sympathectomy, or NT and sgADRB2 tumors of AH43a and AH45b at week 3. Scale bar: 50 μm. Data are either representative of or pooled from at least two independent experiments with n from 3 to 25 (A–C, E–G, I, J, L, and N) or one experiment (D). P values were determined by Student’s t test (A–G, J, and N), one-way ANOVA, and Fisher’s LSD test (I and L). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.
Sympathetic nerve ablation slows ACC growth in vivo. (A–D) AH43a and AH45b tumor volumes from mice treated with 6-OHDA, cervical sympathectomy, propranolol, or sgADRB2 knockdown and NT control at week 3. (E–G) NE concentrations in AH43a and AH45b tumors treated with 6-OHDA, cervical sympathectomy, and propranolol. (H–J) TH+ nerves in AH43a and AH45b tumors and quantification of the percentage TH+ nerves out of all Tubb3+ nerves following propranolol, 6-OHDA, cervical sympathectomy, and sgADRB2 and NT control tumors of AH43a and AH45b at week 3. Scale bar: 50 μm. (K–N) Immunostaining and quantification of CD31+ microvessel length per DAPI+ area in AH43a and AH45b tumors treated with propranolol, 6-OHDA, cervical sympathectomy, or NT and sgADRB2 tumors of AH43a and AH45b at week 3. Scale bar: 50 μm. Data are either representative of or pooled from at least two independent experiments with n from 3 to 25 (A–C, E–G, I, J, L, and N) or one experiment (D). P values were determined by Student’s t test (A–G, J, and N), one-way ANOVA, and Fisher’s LSD test (I and L). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.

Effects of β2R knockdown in AH43a and AH45b. (A) RT-PCR of β2R expression in NT control and ADRB2 KO (sgADRB2) in AH43a and AH45b, using β-actin as endogenous control. (B) MTT assay of NT control and ADRB2 KO (sgADRB2) in AH43a (n = 6) and AH45b (n = 12) treated with isoproterenol. Data are representative of two independent experiments. P values were determined by Student’s t test. *P < 0.5, **P < 0.01, ns: not significant.
Effects of β2R knockdown in AH43a and AH45b. (A) RT-PCR of β2R expression in NT control and ADRB2 KO (sgADRB2) in AH43a and AH45b, using β-actin as endogenous control. (B) MTT assay of NT control and ADRB2 KO (sgADRB2) in AH43a (n = 6) and AH45b (n = 12) treated with isoproterenol. Data are representative of two independent experiments. P values were determined by Student’s t test. *P < 0.5, **P < 0.01, ns: not significant.
Sympathetic ablation decreases ACC NE and TH+ innervation
Following sympathetic ablation, NE concentrations were significantly decreased in both AH43a and AH45b tumors treated with either 6-OHDA (Fig. 4 E) or surgical sympathectomy (Fig. 4 F) as compared with control tumors (P < 0.01 for all comparisons). There was no difference in tumor NE concentration between the control and β2R-blocked tumors (Fig. 4 G). Surgical sympathectomy led to the greatest reduction in tumor NE concentration.
We examined how sympathetic ablation impacts intratumoral innervation using Tubb3 to assess all nerve types and TH to assess sympathetic nerves (Fig. 4 H). The percentage of TH+ nerves was calculated by dividing the area of TH+ nerves by the area of Tubb3+ nerves. We observed a decreased percentage of sympathetic nerves innervating the 6-OHDA-treated and surgically sympathectomized groups as compared with control tumors (Fig. 4 I, P < 0.05 for both comparisons for AH43a, P < 0.001 for both comparisons for AH45b). Surgical sympathectomy led to the lowest TH+ nerve percentages. There were no significant differences in the percentage of TH+ nerves between the propranolol-treated tumors and control tumors (Fig. 4 I). Similarly, in β2R knockdown AH43a and AH45b tumors, there were no significant differences in the TH+ nerve percentages as compared with the NT controls (Fig. 4 J).
Sympathetic ablation decreases ACC PDX microvessel density
Adrenergic innervation in prostate cancer has been linked to tumor angiogenesis (Zahalka et al., 2017). We assessed CD31 staining patterns in ACC PDX with or without sympathetic blockade to determine whether the sympathetic innervation has an indirect effect on cancer growth by altering angiogenesis. Similar to the TH+ nerve distribution pattern, 6-OHDA and surgically sympathectomized tumors showed decreased CD31 signal as compared with control tumors (Fig. 4 K). CD31+ microvessel density was lower in 6-OHDA and surgical sympathectomized tumors (Fig. 4 L, P < 0.05 for AH43a, P < 0.01 for AH45b) as compared with the control tumors. There was no significant difference in CD31+ microvessel density between propranolol-treated tumors and control tumors (Fig. 4, K and L). In AH43a and AH45b sgADRB2 tumors, there was also no significant difference in CD31+ microvessel density as compared with NT control tumors (Fig. 4, M and N). In both normal salivary gland tissue and ACC tumors, CD31+ capillaries are found in close proximity with TH+ nerve fibers.
Inhibition of parasympathetic innervation does not alter ACC PDX growth
We identified VIP+ parasympathetic nerves in ACC specimens (Fig. 1, A–C) and cholinergic muscarinic receptors, with 1, 4, and 5 most expressed in AH43a and AH45b by RT-PCR (Fig. S5 A). Carbachol, an acetylcholine receptor agonist, was added to AH43a and AH45b cells in vitro at varying concentrations and did not affect cell proliferation (Fig. S5 B). Scopolamine, a nonselective muscarinic acetylcholine inhibitor, was used to block parasympathetic innervation to ACC PDX tumors in vivo. There were no significant differences for AH43a or AH45a tumor volumes in scopolamine-treated mice as compared with controls for tumors implanted in the SMG, PG, or flank (Fig. S5 C).

Parasympathetic activity does not affect ACC proliferation or correlate with survival. (A) RT-PCR of relative cholinergic muscarinic receptor expression of AH43a and AH45b. (B) MTT assay of AH43a and AH45b cells treated with carbachol for 24 h or control (n = 6 per group). (C) Tumor volumes at week 3 of AH45b PDX grown in SMG, PG, and flank, treated with scopolamine (n = 15) or control (n = 14). (D) Kaplan–Meier survival curves stratified by parasympathetic neuronal gene expression signature IPAS scores. Parasympathetic signatures do not show significant differences in survival between the high and low groups, stratified by the lowest P value. Parasympathetic neuron shows a nonsignificant trend using the log-rank (Mantel–Cox) test. Data are representative of two independent experiments (B) or one experiment (C). P values were determined by one-way ANOVA, Fisher’s LSD test (B), Student’s t test (C), or log-rank (Mantel–Cox) test (D). ns: not significant.
Parasympathetic activity does not affect ACC proliferation or correlate with survival. (A) RT-PCR of relative cholinergic muscarinic receptor expression of AH43a and AH45b. (B) MTT assay of AH43a and AH45b cells treated with carbachol for 24 h or control (n = 6 per group). (C) Tumor volumes at week 3 of AH45b PDX grown in SMG, PG, and flank, treated with scopolamine (n = 15) or control (n = 14). (D) Kaplan–Meier survival curves stratified by parasympathetic neuronal gene expression signature IPAS scores. Parasympathetic signatures do not show significant differences in survival between the high and low groups, stratified by the lowest P value. Parasympathetic neuron shows a nonsignificant trend using the log-rank (Mantel–Cox) test. Data are representative of two independent experiments (B) or one experiment (C). P values were determined by one-way ANOVA, Fisher’s LSD test (B), Student’s t test (C), or log-rank (Mantel–Cox) test (D). ns: not significant.
Increased adrenergic gene expression signatures correlate with worse survival in ACC patients and with primary site disease in the salivary gland
We analyzed two gene expression datasets (Frerich et al., 2017; Linxweiler et al., 2020) to assess the clinical significance of sympathetic innervation in ACC patients. Adrenergic pathway signatures were obtained from the Gene Ontology and Tabula Sapiens databases. An inferred pathway activation/suppression (IPAS) score was calculated for each patient’s gene expression signature as previously described (Deborde et al., 2022). We performed survival correlations and identified cutoffs based on optimal P values.
Sympathetic nerve gene expression (IPAS) scores were correlated with ACC patient survival. We identified significantly worse survival for ACC patients with higher IPAS scores for the following signatures: sympathetic noradrenergic neuron (hazard ratio [HR]: 2.095 [95% confidence interval [CI]: 0.8505 to 5.159], P = 0.0372), peripheral neuron (HR: 2.182 [95% CI: 1.110 to 4.290], P = 0.0467), noradrenergic neuron development (HR: 2.957 [95% CI: 1.363 to 6.411], P = 0.0002), noradrenergic neuron differentiation (HR: 2.172 [95% CI: 1.145 to 4.120], P = 0.0128), and NE secretion (HR: 2.319 [95% CI: 1.189 to 4.522], P = 0.0068), with a trend for adrenergic neuron (HR: 1.950 [95% CI: 1.012 to 3.758], P = 0.0667) (Fig. 5 A). Further analysis of each pathway showed no significant differences in sex, age, or tumor stage between the high and low groups for any of these signatures, except for tumor stage in the adrenergic neuron signature (Tables S1 and S2). We also examined three parasympathetic-related pathways but did not identify a significant survival separation between high- and low-expressing signatures (Fig. S5 D).

Correlations of sympathetic nerve pathway gene expression signatures with survival for salivary ACC and myoepithelial carcinoma. (A) Kaplan–Meier survival curves using sympathetic nerve gene expression signature IPAS scores. High- and low-expression groups are stratified by lowest P value. Log-rank (Mantel–Cox) test (n = 54). (B) Heatmap of IPAS scores generated for neuronal pathway gene expression signatures for primary salivary ACC (sorted based on TNM stage), metastatic ACC, and primary myoepithelial carcinoma. (C) IPAS score heatmap generated for neuronal pathway gene expression signatures for two ACC patients with synchronous primary and metastatic lesions. (D) Quantification of large-caliber TH+ nerve density in primary salivary ACC (n = 12) and in lung metastases (n = 6) (Student’s t test, *P < 0.05). (E) Left: Single-cell RNA-seq data of ACC cells. UMAPs of primary malignant ACC cells showing the expression level of gene signatures scores for NE secretion, NE transport, and β2R downstream pathway. Right: Boxplot of pathway scores of each primary ACC cell (horizontal lines denote median values, and boxes extend from the first and third quartiles; vertical extending lines denote adjacent values). (F) A scatterplot of single-cell RNA-seq data of ACC cells shows a significant, positive correlation between NE transport and β2R downstream pathway gene expression in seven patients (Pearson’s correlation, R = 0.84, P = 0.017).
Correlations of sympathetic nerve pathway gene expression signatures with survival for salivary ACC and myoepithelial carcinoma. (A) Kaplan–Meier survival curves using sympathetic nerve gene expression signature IPAS scores. High- and low-expression groups are stratified by lowest P value. Log-rank (Mantel–Cox) test (n = 54). (B) Heatmap of IPAS scores generated for neuronal pathway gene expression signatures for primary salivary ACC (sorted based on TNM stage), metastatic ACC, and primary myoepithelial carcinoma. (C) IPAS score heatmap generated for neuronal pathway gene expression signatures for two ACC patients with synchronous primary and metastatic lesions. (D) Quantification of large-caliber TH+ nerve density in primary salivary ACC (n = 12) and in lung metastases (n = 6) (Student’s t test, *P < 0.05). (E) Left: Single-cell RNA-seq data of ACC cells. UMAPs of primary malignant ACC cells showing the expression level of gene signatures scores for NE secretion, NE transport, and β2R downstream pathway. Right: Boxplot of pathway scores of each primary ACC cell (horizontal lines denote median values, and boxes extend from the first and third quartiles; vertical extending lines denote adjacent values). (F) A scatterplot of single-cell RNA-seq data of ACC cells shows a significant, positive correlation between NE transport and β2R downstream pathway gene expression in seven patients (Pearson’s correlation, R = 0.84, P = 0.017).
We also compared adrenergic transcriptomic signatures from primary versus lung metastatic ACC tumors to assess the role of the tumor microenvironment in sympathetic innervation. A heatmap of IPAS scores for the various neural pathway gene expression signatures, including noradrenergic neuron development/differentiation, shows higher expression for primary ACC as compared with metastatic foci and myoepithelial carcinoma (Fig. 5 B). Two patients with ACC had tumor specimens and RNA-seq gene expression data from both synchronous primary and metastatic sites. These matched metastatic sites also demonstrate lower IPAS scores, suggesting that the salivary gland microenvironment more strongly supports neuronal gene expression than the lung (Fig. 5 C). The number of intratumoral, large-caliber TH+ nerves is significantly higher in primary salivary gland ACC as compared with lung metastases (unmatched cases, Fig. 5 D, P < 0.05). This finding is consistent with the absence of TH+ nerves in AH43a and AH45b tumors implanted in the lung (Fig. 3 H).
Analysis of gene expression of primary malignant ACC cells from a published ACC single-cell RNA-seq database (Parikh et al., 2022) showed that the average expression of NE transport gene expression pathway was higher than that of NE secretion. Moreover, known β2R activation–related downstream genes also had a high average expression in ACC cells (Fig. 5 E). Mean expression of NE transport and β2R downstream pathways scores of primary ACC cells in seven patients showed a significant positive correlation (Fig. 5 F, R = 0.84, P = 0.017), suggesting downstream physiologic effects of NE in ACC. These findings suggest that NE secretion by sympathetic nerves acts on ACC cells through β2R signaling.
Discussion
ACC is an aggressive type of salivary carcinoma with a well-described propensity to invade nerves, defined as PNI, which is widely recognized as one of its hallmark characteristics (Amit et al., 2015; Rodriguez-Russo et al., 2021). We show here that ACC progression is supported by axonogenesis and sympathetic innervation. Previous studies have shown that autonomic or sensory nerves support the growth of prostate, gastric, and pancreatic cancers (Magnon et al., 2013; Renz et al., 2018; Saloman et al., 2016; Zhao et al., 2014). The nerves identified in these cancers play a direct role in enabling cancer progression. We speculated that the frequent identification of nerves within ACC may reflect axonogenesis arising from the microenvironment rather than PNI, as commonly interpreted on pathologic diagnosis. PNI involves cancer cell transit along nerves and is supported by the release of ligands from nerves to attract cancer cells, macrophage recruitment, and Schwann cell reprogramming (Bakst et al., 2017; Deborde et al., 2016, 2022; He et al., 2014, 2015). Because the autonomic nervous system controls salivary gland development and physiological function (Kinjo et al., 1983; Nedvetsky et al., 2014), we hypothesized that autonomic nerves might directly influence salivary carcinoma growth rather than solely serving as a conduit for PNI.
Studying ACC is challenging due to its rarity, lack of cell lines, and few animal models available. We examined the role of the autonomic nervous system in salivary ACC using patient surgical specimens, bulk and single-cell gene expression transcriptomic datasets, and two new orthotopic ACC PDX murine models. These orthotopic ACC PDX tumors recapitulate the original patient specimen’s morphology and exhibited characteristics of PNI. The orthotopic location provides an environment similar to that of their origin and better suited for studying the tumor microenvironment (Byrne et al., 2017).
We provide evidence supporting the identification of axonogenesis in ACC. IF microscopy of ACC reveals small-caliber TH+ nerves growing in a disorganized pattern of single nerve fibers that arise near the periphery of the tumor or adjacent to large-caliber TH+ nerves. This configuration is reminiscent of axonogenesis that occurs during development or neural repair. Ayala et al. (2008) used PGP9.5 to identify “asymmetric” cancer-related axonogenesis in prostate cancer, while Madeo et al. (2018) employed βIII tubulin to detect nerve “twigs” in head and neck cancer (Ayala et al., 2008; Madeo et al., 2018). Guillot et al. (2022) used TH staining to identify sympathetic “sprouts” in a KIC mouse model of pancreatic cancer, while Baloh et al. (2022) utilized NFH staining to identify “disorganized and regenerative axons” in ALS models (Baloh et al., 2022; Guillot et al., 2022). We also note that these disorganized single nerve fibers only appear within tumor tissue and not adjacent normal salivary gland tissue, suggesting that axonogenesis is induced by the cancer cells or other components of the tumor microenvironment.
The increased nerve density observed in human ACC specimens may reflect a long-term remodeling process from tumor-nerve interactions that require an extended period of time. In contrast, in the PDX model, most intratumoral nerves are located near the junction of normal salivary tissue and ACC. This pattern suggests that these nerves may have originated from neural structures in the adjacent normal murine salivary gland and grown into the tumor within the brief experimental timeframe. The short duration of the PDX model may not provide sufficient time for substantial neo-axonogenesis to occur within the tumor microenvironment. ACC PDXs in mouse salivary glands show faster growth as compared with the flank, highlighting the importance of the microenvironment. Small differences between ACC growth in the SMG and PG might be attributed to differences in gland size, serous vs. mucous cell composition, and innervation patterns between the two gland types.
A clear distinction between definitions of PNI and tumor innervation remains undefined in the literature (Silverman et al., 2021). We hypothesize that ACC innervation is mistaken as PNI, defined pathologically when tumor cells are identified inside or around nerves seen on H&E staining. H&E staining cannot resolve single nerve fibers, which requires IF microscopy with antibodies targeting specific neural proteins and is not performed during routine pathologic diagnosis. We show that CM from ACC stimulated SG neurite outgrowth and speculate that there may be a continuum between PNI and tumor innervation where both processes are at play. Studies have highlighted the role of nerve growth factor (NGF) in ACC progression and its association with PNI (Hao et al., 2010) and poor prognosis (Kobayashi et al., 2015). NGF expression is upregulated in ACC as compared with normal salivary gland (Tang et al., 2023) and promotes tumor cell migration along nerve fibers through the tyrosine kinase A receptor (Wang et al., 2006). NGF has also been identified in tumor-derived exosomes, facilitating nerve infiltration (Xu et al., 2019). The proliferation of sympathetic nerve fibers we demonstrate in ACC might be related to NGF activity.
The sympathetic nervous system affects tumor initiation, progression, metastasis, angiogenesis, and creates an immunosuppressive microenvironment (Cole et al., 2015; Faulkner et al., 2019; Gysler and Drapkin, 2021). The sympathetic nervous system and NE regulate tumor growth and vascularization through cAMP-PKA signaling, promoting the expression of VEGF, MMP2, and MMP9 (Thaker et al., 2006). In addition, sympathetic innervation of endothelial cells drives an angio-metabolic switch that supports prostate cancer growth (Zahalka et al., 2017). Although we did not observe an impact of sympathetic blockade on microvessel density in our PDX model, this might be the related to timing and duration of the blockade in these models. Inhibition of endothelial cell proliferation with propranolol has shown a wide range of responses (Chen et al., 2012; Lee et al., 2009; Maccari et al., 2017; Thaker et al., 2006). In our PDX model, the continued presence of NE during propranolol treatment may have limited its observable impact on microvessel density.
Stress-induced sympathetic activation contributes to an immunosuppressive tumor microenvironment by driving M2 macrophage infiltration (Sloan et al., 2010), T cell exhaustion (Globig et al., 2023; Qiao et al., 2021), and blocking β-adrenergic signaling decreases PD-1 expression in CD8+ T cells (Bucsek et al., 2017). β-Adrenergic receptors play key roles in cancer progression and tumor immunity (Cole and Sood, 2012). β2R is widely implicated in enhancing cancer cell proliferation, migration, and invasion (Jeong et al., 2022). β2R also modulates immune cell function, suppressing cytotoxic activity and promoting an immunosuppressive microenvironment (Mohammadpour et al., 2019). Chronic stress, through β-adrenergic receptor activation, suppresses CD8+ T cell function, impairing anti-tumor immunity and immunotherapy efficacy (Bucsek et al., 2017). Sympathetic input also contributes to an unfavorable microenvironment for immunotherapy (Linxweiler et al., 2020). Combining β-blockers with immune checkpoint blockades might overcome such an immunosuppressive environment (Mellgard et al., 2023), and targeting βARs is one emerging strategy to enhance anti-tumor immunity (Fjæstad et al., 2022; Shaashua et al., 2017).
We show here that sympathetic nerves innervate ACC to support tumor growth, with NE contributing to direct effects on cancer cell proliferation. Sympathetic blockade has previously been shown to correlate with improvement in clinical outcomes in breast, prostate, and pancreatic cancer (Grytli et al., 2014; Melhem-Bertrandt et al., 2011; Na et al., 2018). The inhibition of ACC sympathetic innervation through widely available drugs such as β-adrenergic blockers merits further investigation in clinical trials.
In 2020, Amit et al. (2020) showed decreased oral squamous cancer progression with sensory denervation or pharmacologic blockade of adrenergic receptors. Transcriptomic data also revealed an adrenergic differentiation signature. However, chemical sympathectomy did not impact cancer progression. The authors suggest that sensory nerves transdifferentiate into adrenergic nerves to promote oral cancer progression and that this is driven by loss of p53. We demonstrate here that adrenergic stimulation also promotes ACC progression, and any type of adrenergic inhibition (adrenergic receptor blockade, surgical or chemical sympathectomy) inhibits ACC progression.
We identified significantly worse survival for ACC patients with higher signature scores for sympathetic noradrenergic neuron, peripheral neuron, noradrenergic neuron development, noradrenergic neuron differentiation, and NE secretion. The identification of neural developmental pathways suggests that the mechanism of ACC innervation is similar to the molecular pathways used during embryologic nerve development (Scott-Solomon et al., 2021). Single-cell RNA-seq data from ACC cells showed expression of genes involved in NE transport and genes downstream of the β2R. These two pathways are significantly and positively correlated, suggesting that NE has physiologic effects in ACC cells. In contrast, NE secretion pathways were minimally expressed by ACC cells, consistent with NE being released from the sympathetic nerves. Identifying neuronal cells in single-cell RNA-seq may be challenging since the neuronal cell body may be distant from the tumor and the low content of RNA in axons. Single-nuclei RNA-seq to maximize the collection of neuronal cell transcripts should be considered for future analysis.
The survival differences in selected neuronal pathways are independent of tumor stage. This suggests that ACC tumor innervation may not develop linearly with tumor stage. Similarly, the survival differences are also independent of distant metastasis in colorectal (Albo et al., 2011) and pancreatic cancers (Ferdoushi et al., 2021). In contrast, positive correlations between nerve density and tumor stage were found in liver (Zhang et al., 2017) and gastric cancers (Zhao et al., 2014). These correlations of nerve densities did not distinguish the subtype of nerve in the tumors.
In summary, we demonstrate an intimate developmental relationship between ACC and the sympathetic nervous system. This unique relationship between ACC and sympathetic nerves does not simply represent PNI but rather new tumor axonogenesis and innervation that drive tumor growth. These findings reveal new opportunities for sympathetic blocking agents to improve available therapies for ACC, which currently have limited efficacy.
Materials and methods
ACC PDX models and cell culture
Tumor tissue from two patients with ACC was collected under an approved Institutional Review Board protocol (protocol #14–091). The tissue was immediately minced, mixed (1:1) with Matrigel (Corning), and implanted subcutaneously in 6–8-wk-old female NSG mice (#005557; Jackson Laboratory; Strain) to generate PDX as previously described (Mattar et al., 2018). Mice were cared for following guidelines approved by the Memorial Sloan Kettering Cancer Center Institutional Animal Care and Use Committee (IACUC) and Research Animal Resource Center under an approved IACUC protocol (protocol # 04-03–009).
Established PDX tumors were minced and digested in GentleMACS C tube (130-093–237; Miltenyl Biotec) with 3 ml of digestion buffer, consisting of HBSS++, collagenase IV (400 U/ml) (CLS-4; Worthington Biochemical Corp.), dispase II (2 U/ml) (4942078001; Sigma-Aldrich), and DNase I (200 U/ml) (LS002139; Worthington Biochemical Corp.), using a GentleMACS Octo dissociator with heater (130-096–427; Miltenyl Biotec). The resulting cell suspension was filtered through a Falcon 70-μm cell strainer (352350; Corning), followed by three consecutive washing steps in HBSS++ with 1% BSA (BP-1600-100; Thermo Fisher Scientific).
AH43a cells were grown in DMEM, high glucose (DMEM HG): Ham’s F12 nutrient mixture (F12) (1:1) with 10% FBS, nonessential amino acids, 2 mM L-glutamine, penicillin, and streptomycin. AH45b cells were grown in 3T3-J2 (EF3003; Kerafast) CM supplemented with hydrocortisone (0.4 μg/ml) (74142; Stem Cell), insulin–transferrin–selenium (5 μg/ml) (41400045; Gibco), cholera toxin (8.6 ng/ml) (C8052; Sigma-Aldrich), human epidermal growth factor recombinant protein (10 ng/ml) (PHG0315; Gibco), and ROCK-inhibitor Y27632 (10 μM) (S1049; SelleckChem) (Panaccione et al., 2017). 3T3-J2 cells were cultured in T-225 flasks for 3–4 days in 45 ml of DMEM HG, supplemented with 10% HyClone iron-supplemented calf serum (SH30072.03; Cytiva), penicillin, and streptomycin. The CM was then filtered through a 0.22-μm filter (596–3320; Nalgene) and stored at 4°C.
To generate CRISPR knockdown of β2R, AH43a and AH45b were transfected with lentiCas9-EGFP (#63592; Addgene) (Chen et al., 2015), followed by guide RNA cloned into pU6-sgRNA EF1apuro-T2A-BFP (#60955; Addgene) (Gilbert et al., 2014). Transfected cells were selected using flow cytometry for EGFP and BFP double-positive cells, and total RNA was extracted for RT-PCR to confirm gene knockout.
Animal experiments
All mouse procedures were done following IACUC guidelines and an approved protocol (#05-04-006) at the Memorial Sloan Kettering Cancer Center. NSG, B6N.FVB-Tg(Mpz-cre)26Mes/J (#017927), and B6.Cg-Gt(ROSA)26Sortm14(CAG-tdTomato)Hze/J (#007914) mice were all from Jackson Laboratory. P0-Cre mice were crossed with Rosa26tdTomato to create P0TdTomato, then crossed with NSG mice to obtain NSG.B6-P0tdTomato to generate immunocompromised red fluorescent Schwann cells expressing mice. 6–8-wk-old mice were used in all animal experiments.
Dual fluorescent and bioluminescent human ACC, 5 × 104 AH43a and AH45b cells, suspended in 1–5 μl of Matrigel, were injected orthotopically to the SMG or PG or subcutaneously to the flank. Tumor volumes were accessed weekly with Vevo 2100 imaging system (FUJIFILM VisualSonics Inc.) and/or IVIS spectrum in vivo imaging system (PerkinElmer). A final tumor volume measurement was taken at the time of the necropsy in 3 wk.
AH43a and AH45b, 5 × 104 cells, were injected orthotopically to the left SMG and treated with intraperitoneal injection of 6-OHDA (H4381; Sigma-Aldrich) at 100 mg/kg on the day of tumor injection and 250 mg/kg 48 h later. 6-OHDA was freshly prepared in 0.1% ascorbic acid (A2218; Sigma-Aldrich) and diluted in saline (306546; BD) to a total volume of 200 μl. Propranolol (P0884; Sigma-Aldrich) was dissolved in water, diluted in saline, and injected intraperitoneally daily, 5 days/week, at 10 mg/kg. Additional propranolol was added to the drinking water at 500 mg/liter and replaced weekly to ensure continuous exposure.
Bilateral cervical sympathectomy was performed by medially exposing and removing the cervical SG (Wang et al., 2022). Sham surgery was performed as a control by exposing but without removing the SG. A successful ganglionectomy is confirmed by observing Horner’s syndrome (ptosis) after the surgery. 5 × 104 cells of HTB-41 were injected orthotopically to the right SMG and treated with either sham surgery or sympathectomy.
Scopolamine (S0929; Sigma-Aldrich), a competitive nonselective muscarinic acetylcholine antagonist, was given intraperitoneally daily at a concentration of 1 mg/kg in a total volume of 200 μl of NaCl in NSG mice injected with 5 × 104 cells of AH45b into the left SMG, right PG, and right flank.
AH43a and AH45b, with sgADRB2 knockdown or NT control, were injected (5 × 104 cells) to the left SMG of NSG mice. Tumor volume was measured 3 wk after injection. Dual fluorescent and bioluminescent AH43a and AH45b, 5 × 105 to 1 × 106 cells in 100 μl PBS, were injected through the tail vein to NSG mice and followed weekly to confirm tumor colonization in the lungs.
PDX tissue preparation for IF staining and NE quantification
Three weeks after AH43a and AH45b injection, mice were euthanized and transcardially perfused with heparin (20 U/ml, 306515; BD) and 4% paraformaldehyde (15714; Electron Microscopy Science). Harvested tumors were frozen in milestone cryoembedding compound (6703MILE01; Milestone Medical) for frozen sections at 5- or 40-μm thickness. Fresh tumors or 500-μm–thick frozen slices were fixed in 10% buffered formalin (SF-100; Thermo Fisher Scientific), processed in a tissue processor (Leica), and embedded in paraffin blocks. Deparaffinization and heat-induced antigen retrieval with 10 mM sodium citrate buffer was done on 5–20-μm paraffin sections before immunofluorescent staining. Sections were blocked in 10% normal goat serum (S26LITER; Millipore) with 1% BSA, 0.1% Triton-X (BP151–500; Thermo Fisher Scientific), and 0.05% Tween20 (BP337–500; Thermo Fisher Scientific) for 1 h at room temperature. Primary antibodies were prepared in 3% BSA with 0.1% Triton-X in PBS and incubated with sections overnight at 4°, followed by Hoechst (H1399; Thermo Fisher Scientific) nuclear stain and corresponding secondary antibodies for 1 h at room temperature before sections were mounted in Mowiol solution (81381; Sigma-Aldrich). AH43a or AH45b cells (5 × 105) were seeded in 35-mm glass bottom dish (P35G-1.5–20-C; MatTek) overnight. Cells were rinsed with PBS and fixed in 2% paraformaldehyde for 20 min at room temperature. After membrane permeabilization with 0.1% Triton-X, cells were blocked and incubated with primary and secondary antibodies, as described above. Mouse cervical SG were used as positive controls for the TH immunostaining. Mouse vagus nerve and parasympathetic ganglia in the mouse SMG were used as positive controls for the VIP immunostaining.
Stained sections were digitized with Pannoramic 250 Flash or Pannoramic confocal scanners (3DHistech) and viewed with CaseViewer software (3DHistech). Images were analyzed and quantified using ImageJ/Fiji (National Institutes of Health), where thresholding was used to isolate the fluorescent signal and measure the area and intensity. Masks were created, and an image calculator was used to measure the overlapping area and intensity between two signals. TH+ fluorescent nerves were identified and counted from a single section of various human salivary gland tumor specimens using IF and corresponding H&E images. TH+ and NFH+ nerve areas were quantified and divided by the tumor area (mm2) to normalize data for each tumor. Overlay images of IF and H&E were created using Photoshop (Adobe) using the same section sequentially stained with IF and H&E. Large-caliber nerves from both images were used as a point of reference to guide the overlay positioning. For quantification of TH+ nerves in a panel of salivary neoplasms, a blinded observer was used for validation.
For PDX sympathetic nerve quantification, TH+ fluorescent areas were divided by areas of a pan-neuronal marker (representing all nerves) and expressed as the percentage of TH+ nerves out of all nerves. To confirm neuronal identity of IF signals, all samples were stained with pan-neuronal markers, including NFH, Tubb3, and corresponding H&E images. For microvessel quantification, CD31+ areas were skeletonized using ImageJ plug-in and transformed to an estimated average length of the longest branch based on the parameters used in scanning the slides and normalized to the total area selected.
Fresh tumors from each treatment were homogenized using GentleMACS M tube in 1 ml of buffer consisting of 0.01 N HCl, 1 mM EDTA, and 4 mM sodium metabisulfite. The NE concentration was quantified using a noradrenaline research ELISA kit (BA E-5200; Labor Diagnostika Nord).
Patient specimens
Human salivary tumor specimens were obtained for immunostaining, including nine ACC, three polymorphous adenocarcinomas, five mucoepidermoid carcinomas, three myoepithelial carcinomas, eight acinic cell carcinomas, five pleomorphic adenomas, and an ACC tissue microarray (Ho et al., 2013). Nine additional ACC specimens (three primary and six metastatic) from a previous report (Linxweiler et al., 2020) were also included. Patient studies were conducted in compliance with the ethical guidelines of the Belmont Report and through a Memorial Sloan Kettering Institutional Review Board–approved protocol (#15–052). Patient specimens were sectioned into 5-μm sections and processed as above.
MTT assay
AH43a and AH45b, 3 × 103 cells/well, were plated in 96-well plates (353910; Corning) in complete growth media overnight. After cells were attached to the plates, cells were starved in serum-free media for 24 h. NE (A7256; Sigma-Aldrich), prepared in 100 μM ascorbic acid, or propranolol or carbachol, (C4382; Sigma-Aldrich) was added in 100 μl/well serum-free media to each designated well for another 24 h. For NE and propranolol combination treatment, cells were pretreated with propranolol for 1 h before adding the drug combination. MTT solution (5 mg/ml, 10 μl/well) (M2128; Sigma-Aldrich) was added at the end of the treatment. After 4 h of incubation, 100 μl of isopropanol (BP2618; Thermo Fisher Scientific) with 0.04N HCl (1090571000; Sigma-Aldrich) was mixed in each well. The plate was read by The Synergy HT plate reader (Biotek) at 570 nm, reference 630 nm.
AH43a and AH45b with sgADRB2 or NT control, 3 × 103 cells/well, were plated in 96-well plates with serum starvation after attached to the plate as mentioned above. Serum-free media with or without isoproterenol (I5627; Sigma-Aldrich) were added to each designated well for 24 h, and cell viability assay was performed as above.
SG co-culture
Cervical and thoracic SG were harvested from NSG or NSG.B6-P0tdTomato mice using a bright-field (SZ61 Olympus) or fluorescent (Zeiss) stereomicroscope. 25 to 50 cells of AH43a or AH45b were suspended in 5 μl of Matrigel before explanting the harvested SG in 35-mm glass bottom dish. Co-cultures were grown in DMEM HG with 1 mM sodium pyruvate, 1.5 g/liter sodium bicarbonate, and 10% FCS for 7–10 days. Tumor spheres were imaged using Axio Observer Z1 (Zeiss), and the number of spheres was counted manually. Tumor spheres and SG neurites interfaces were imaged with a Leica SP8 laser scanning confocal microscope (Leica microsystems), using 20×/0.75 water immersion objective imaged at 37°C in culture media. Images were obtained and processed with Leica Application Suite X software.
SG explants provide a continuous source of NE, which has limited stability in culture. SG explants with or without adding AH43a or AH45b were cultured with or without 10 μM propranolol for 7 days. CM was harvested from SG explants and added to wells with AH43a or AH45b spheroids in Matrigel. The number of spheroids was counted and presented as percent of control.
Additionally, CM from cultured AH43a cells was collected and filtered through a 0.22-μm filter (596–3320; Nalgene) before adding to wells with SG only. SG were imaged using Axio Observer Z1 microscope. To quantify the neurites covered area using ImageJ/Fiji, a variance filter was applied to the image to segment the cells based on contrast. A mask was created, and the particles were analyzed to determine the total area and the largest contiguous piece using ImageJ.
Western blot
Protein lysates were collected from 1 × 105 cells/well of AH43a and AH45b plated in 24-well plates (353047; Corning) and grown in complete growth media overnight and then starved in 0.1% FCS media for 24 h, followed by NE, propranolol, and combination treatments, same as MTT assays. Cells were rinsed with PBS and lysed in 50 μl of Laemmli SDS buffer (J60660-AC; Thermo Fisher Scientific) with sample reducing agent (NP0009; Thermo Fisher Scientific) and denatured for 20 min at 95°C. An equal amount of samples was loaded to 4–15% gel (456–1086; Bio-Rad) and electrophoresed at 100 V. Protein samples were then transferred onto a polyvinylidene fluoride membrane (170–4273; Bio-Rad) using Trans-Blot Turbo Transfer System (Bio-Rad). Membranes were blocked in 5% nonfat milk (170–6404; Bio-Rad) and incubated with primary and secondary antibodies prepared in TBST buffer (T9511; Teknova) with 5% BSA. Images were taken with Odyssey DLx imager (Licor) and analyzed with LI-COR image studio software (Licor).
Characterizing sympathetic-related pathways in bulk and single-cell RNA-seq of ACC
We obtained two bulk ACC RNA-seq databases from previous publications (Frerich et al., 2017; Linxweiler et al., 2020). An IPAS score (Deborde et al., 2022) was calculated for each gene expression pathway and used to generate heatmaps using Morpheus (https://software.broadinstitute.org/morpheus). All cases were included in all analyses, except in a subset of TNM analysis where three cases were excluded because of missing tumor stage information. To focus on sympathetic-related pathways, well established gene signatures and sympathetic/parasympathetic related pathways were selected from Tabula Sapiens and Gene Ontology biological process. Cases were ranked by ascending IPAS scores and separated into high and low groups determined by using Kaplan–Meier method and log-rank test to identify the lowest P value and to assess survival differences between groups. Pearson’s Chi-squared test, Wilcoxon rank-sum test, and Fisher’s exact test were used to assess for differences in gender, age, and TNM stage (stage I–III versus IV) between high and low groups in each selected pathway.
For single-cell RNA-seq gene expression pathway scores, each pathway’s gene expression was calculated using average log2(TPM+1) values in primary ACC cells from a published ACC single-cell RNA-seq database (Parikh et al., 2022). Pathway genes were obtained from GO database, while a β2R pathway consisted of downstream genes related to β2R activation, including RAF1, MAPK1, PRKACA, AKT1, AKT2, AKT3, CREB1, CREBBP, BCL2, BAD, JUN, FOS, and MYC. Pearson correlation across average pathway scores in each patient was used to calculate pathway correlations.
Antibodies
Anti-NFH polypeptide (ab8135), anti-VIP (ab22736), anti-β2 adrenergic receptor (ab182136), anti-CD31 (ab124432), anti-TRPV1 (ab203103), and anti-S100 (ab52642) (Abcam); anti-TH and anti-Tubb3 (801202) (801202; BioLegend); Alexa Fluor 488/568 anti-rabbit and -anti-mouse (A11034, A11036, A11029, and A11031; Invitrogen); anti–phospho-ERK (#9101), anti–phospho-RB (#9308), anti–phospho-CREB (#9198), and anti–phospho-Bcl2 (#2827) (Cell Signaling Technology); anti–phospho-cJun (PA5–17890; Invitrogen); anti-GAPDH (MAB374; Millipore); IRDye800CW goat anti-rabbit (925–32211; LiCor); and IRDye680RD goat anti-mouse (925–68070; Licor).
RT-PCR
Extraction of mRNA from 5 × 106 cells of AH43a, AH45b, and HTB-41 using RNeasy mini kit (74704; Qiagen) was then reverse transcribed into cDNA using qScript cDNA supermix (95048-100; Quantabio). Quantitative PCR was performed with PerfeCTa SYBR Green FastMix Low ROX (95074-012; Quantabio) and ran on a ViiA7 RealTime PCR System (Applied Biosystems).
Primers sequences are ACTB forward, 5′-TGTGATGGTGGGAATGGGTCAG-3′; ACTB reverse, 5′-TTTGATGTCACGCACGATTTCC-3′; Adrb1 forward, 5′-TCCTTGTGTAGGGCAAACCC-3′; Adrb1 reverse, 5′-CGCCTGGTCCTTCCAACTAA-3′; Adrb2 forward, 5′-TGTGGGCCATCAAGGTACTG-3′; Adrb2 reverse, 5′-AAGGATGCCCTTCCTTCTGC-3′; Adrb3 forward, 5′-TACTCTGCGCTGGCTTTTGA-3′; Adrb3 reverse, 5′-AAAGGCTCAAGCTCACTCCC-3′; Chrm1 forward, 5′-TGACCGCTACTTCTCCGTGACT-3′; Chrm1 reverse, 5′-CCAGAGCACAAAGGAAACCA-3′; Chrm2 forward, 5′-TCACAAAACCTCTGACCTACCC-3′; Chrm2 reverse, 5′-TCCACAGTTCTCCACCCTACAA-3′; Chrm3 forward, 5′-ACCATCCCTCAACTCCACCAAGT-3′; Chrm3 reverse, 5′-GGAAAACTGCCTCCATCGTC-3′; Chrm4 forward, 5′-TCGCTATGAGACGGTGGAAA-3′; Chrm4 reverse, 5′-AGCACAACCAATAGCCCAAG-3′; Chrm5 forward, 5′-GAAAGCAGCCCAGACACTGA-3′; Chrm5 reverse, 5′-AGCACAACCAACAGCCCAAG-3′.
Plasmid construction
To generate pLVX-EF1α-IRES-LucZsGreen1, pLVX-EF1α-IRES-Luciferase/EGFP was constructed by cloning IRES-Luciferase/EGFP from pSico-CAG-v-H-Ras-IRES-Luciferase/EGFP (#58959; Addgene) using NotI and MluI restriction sites (forward primer: 5′-AAGGAAAAAAGCGGCCGCCGCCCCTCTCCCTCCCCCCC-3′, reverse primer: 5′-AAACGACGCGTCTACTTGTACAGCTCGTCCATGCCG-3′). The EGFP in the resulting construct was replaced by ZsGreen1 cloned from pLVX-EF1α-IRES-ZsGreen1 (631982; Takara Bio Inc.) using BamHI and MluI sites (forward primer: 5′-CGGGATCCACCGGTCGCCACCATGGCCCAGTCCAAGCAC-3′, reverse primer: 5′-TTCCAGACGCGTTCAGGGCAA-3′). sgRNA sequencing targeting human β2-receptor (ADRB2), 5′-TTGGAACGGCAGCGCCTTCTTGCGTTTAAGAGC-3′, and a nontargeting control, Gal4, 5′-TTGGAACGACTAGTTAGGCGTGTAGTTTAAGAGC-3′, were cloned into pU6-sgRNA EF1a-puro-T2A-BFP (#60955; Addgene) using BstXI and BlpI sites.
Statistical analyses
Statistical analyses were performed using an unpaired two-tailed Student’s t test, one-way ANOVA, Fisher’s least significant difference test, log-rank (Mantel–Cox), and Pearson’s correlations using Prism 9. Statistical significance was defined as P < 0.05. All data are represented as mean ± standard error of the mean.
Online supplemental material
Fig. S1 quantifies the TH+ nerve sprouts in human salivary gland neoplasms, showing high density in ACC and low or absent levels in other tumor types. Fig. S2 demonstrates the reduced intratumoral nerve sprouting and no effect of sympathetic ablation in a non-ACC salivary tumor model. Fig. S3 validates the selective sympathetic ablation of normal murine salivary glands, showing no impact on vascularity or physiological parameters. Fig. S4 examines the effects of β2R knockdown in AH43a and AH45b, including MTT assays of cell proliferation and response to isoproterenol. Fig. S5 shows that parasympathetic activity does not affect ACC proliferation or correlate with survival. Tables S1 and S2 contain 54 ACC patient clinical characteristics and their associations with neuronal gene expression pathways.
Data availability
The source data for Fig. 2 A; Fig. 5, A–C; and Tables S1 and S2 are openly available from the National Center for Biotechnology Information Sequence Read Archive under accession numbers SRP109264, SRP096726, PRJNA601423 (Linxweiler et al., 2020), and PRJNA287156 (Frerich et al., 2017). The source data for Fig. 5, E and F are available at GEO (GSE210171) and dbGaP (phs003070.v1) (Parikh et al., 2022).
Acknowledgments
We thank S.A. Ness for providing RNA-seq data and O. Rukovets for editorial support.
This study was supported, in part, by the National Institutes of Health (NIH)/National Cancer Institute (NCI) grant R01CA219534, NIH/NCI Cancer Center Support Institutional Grant P30CA008748, and by generous funding from Mr. Leerom Segal.
Author contributions: C.-H. Chen: conceptualization, data curation, formal analysis, investigation, methodology, project administration, resources, visualization, and writing—original draft, review, and editing. B. Reva: formal analysis. N. Katabi: data curation and writing—review and editing. A. Wizel: data curation, formal analysis, investigation, software, and visualization. H. Xu: methodology. A.L. Ho: methodology, resources, visualization, and writing—review and editing. L.G.T. Morris: formal analysis and writing—review and editing. R.L. Bakst: conceptualization, data curation, formal analysis, methodology, and writing—original draft. A.S. Parikh: formal analysis, resources, and writing—review and editing. Y. Drier: formal analysis, resources, and supervision. S. Deborde: conceptualization, formal analysis, investigation, methodology, supervision, visualization, and writing—original draft, review, and editing. R.J. Wong: conceptualization, formal analysis, funding acquisition, methodology, project administration, resources, supervision, visualization, and writing—original draft, review, and editing.

