


Considerable evidence suggests that day–night rhythms in the functional expression of subthreshold potassium (K+) channels regulate daily oscillations in the spontaneous firing rates of neurons in the suprachiasmatic nucleus (SCN), the master circadian pacemaker in mammals. The K+ conductance(s) driving these daily rhythms in the repetitive firing rates of SCN neurons, however, have not been identified. To test the hypothesis that subthreshold Kv12.1/Kv12.2-encoded K+ channels play a role, we obtained current-clamp recordings from SCN neurons in slices prepared from adult mice harboring targeted disruptions in the Kcnh8 (Kv12.1−/−) or Kcnh3 (Kv12.2−/−) locus. We found that mean nighttime repetitive firing rates were higher in Kv12.1−/− and Kv12.2−/− than in wild type (WT), SCN neurons. In marked contrast, mean daytime repetitive firing rates were similar in Kv12.1−/−, Kv12.2−/−, and WT SCN neurons, and the day–night difference in mean repetitive firing rates, a hallmark feature of WT SCN neurons, was eliminated in Kv12.1−/− and Kv12.2−/− SCN neurons. Similar results were obtained with in vivo shRNA-mediated acute knockdown of Kv12.1 or Kv12.2 in adult SCN neurons. Voltage-clamp experiments revealed that Kv12-encoded current densities in WT SCN neurons are higher at night than during the day. In addition, the pharmacological block of Kv12-encoded currents increased the mean repetitive firing rate of nighttime, but not daytime, in WT SCN neurons. Dynamic clamp-mediated subtraction of modeled Kv12-encoded currents also selectively increased the mean repetitive firing rates of nighttime WT SCN neurons. Despite the elimination of the nighttime decrease in the mean repetitive firing rates of SCN neurons, however, locomotor (wheel-running) activity remained rhythmic in Kv12.1−/−, Kv12.2−/−, and Kv12.1-targeted shRNA-expressing, and Kv12.2-targeted shRNA-expressing animals.
Introduction
Neurons in the suprachiasmatic nucleus (SCN), the master circadian pacemaker in mammals, including humans, display day–night oscillations in spontaneous repetitive firing rates that drive daily rhythms in physiology and behavior (Reppert and Weaver, 2001; Hastings et al., 2018, 2019; Michel and Meijer, 2020). In vivo recordings from the rat SCN, isolated from synaptic and humoral inputs, were the first to demonstrate cell-autonomous circadian rhythms in the electrical activity of the SCN, with neuronal firing rates higher during the day than at night (Inouye and Kawamura, 1979). Numerous subsequent studies have confirmed that the repetitive firing rates of SCN neurons are higher during the day than at night, with daytime firing rates averaging about 5 Hz and nighttime firing rates averaging about 1 Hz (Kuhlman and McMahon, 2006; Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020). These daily rhythms in repetitive firing rates are accompanied by alterations in input resistances (higher during the day than at night) and membrane potentials (more depolarized during the day than at night), linked to daily changes in subthreshold potassium (K+) conductance(s) (Kuhlman and McMahon, 2006; Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020). An alternative “bicycle” model for the circadian regulation of membrane excitability in the SCN, involving a daytime increase in an inward Na+ leak current (causing membrane depolarization and increased firing) and a nighttime increase in a K+ current (causing membrane hyperpolarization and decreased firing), has also been proposed (Flourakis et al., 2015). The daytime depolarization in the membrane potentials of SCN neurons, however, is associated with increased, not decreased, input resistance, as would result from opening inward Na+ leak channels. Even in the bicycle model (Flourakis et al., 2015), therefore, a reduction in daytime K+ conductance is required. Interestingly, this daily pattern (higher firing rates and input resistances during the day than at night) is observed in both diurnal and nocturnal animals (Nunez et al., 1999; Kumar Jha et al., 2015), suggesting that the underlying molecular mechanism(s) is conserved.
Although the daily oscillations in the membrane potentials and input resistances of SCN neurons indicate that day–night changes in subthreshold K+ conductance(s) are required for rhythmic changes in repetitive firing rates, the critical K+ conductance(s) has not been identified (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020). Several studies have identified roles for voltage-gated K+ (Kv) channels encoded by the Kcnc1 (Kv3.1; Itri et al., 2005; Kudo et al., 2011), Kcnc2 (Kv3.2; Itri et al., 2005; Kudo et al., 2011), Kcna4 (Kv1.4; Granados-Fuentes et al., 2012, 2015), Kcnd1 (Kv4.1; Itri et al., 2010; Hermanstyne et al., 2017), and Kcnd2 (Kv4.2; Itri et al., 2010; Granados-Fuentes et al., 2012, 2015) pore-forming (α) subunits and for large conductance voltage- and Ca2+-dependent K+ (BK) channels (Meredith et al., 2006; Kent and Meredith, 2008; Whitt et al., 2016) in regulating the repetitive firing rates of SCN neurons. Although day–night differences in the densities of several of these K+ currents (Itri et al., 2005, 2010; Meredith et al., 2006; Kudo et al., 2011; Hermanstyne et al., 2017) and in the expression levels of transcripts (http://circadb.hogeneschlab.org) encoded by the underlying K+ channel subunit genes (Itri et al., 2005; Meredith et al., 2006) have been reported, none of these K+ channels has been shown to control the day–night switch in the input resistances, membrane potentials, and spontaneous repetitive firing rates of SCN neurons (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020). The K+ conductance(s) driving the cell-autonomous circadian rhythms in the repetitive firing properties of SCN neurons remain to be identified. Similar to other types of central and peripheral neurons, in situ hybridization and quantitative RNA expression profiling data show that transcripts encoding a number of other K+ channel α subunits are expressed in the SCN (Panda et al., 2002; Harvey et al., 2020). Of particular note are two members of the Ether-à-go-go-like (Elk) subfamily of Kv channel α subunits, Kcnh8 and Kcnh3, which encode the Kv12.1 and Kv12.2 α subunits, respectively, and are predominantly expressed in the brain (Engeland et al., 1998; Miyake et al., 1999; Zou et al., 2003). Heterologous expression studies have demonstrated that Kv12.1 or Kv12.2 generate voltage-gated outward K+ currents that activate at subthreshold membrane potentials (Trudeau et al., 1999; Becchetti et al., 2002; Zou et al., 2003; Zhang et al., 2009, 2010; Li et al., 2015), but display marked differences in inactivation: Kv12.2-encoded currents partially inactivate at depolarized membrane potentials (greater than +20 mV), whereas Kv12.1-encoded outward currents do not (Engeland et al., 1998; Miyake et al., 1999). The voltage-dependences of activation of Kv12.x-encoded K+ currents are affected by Zn2+ and some (but not all) other divalent cations (Zhang et al., 2010) and are modulated by phosphatidylinositol 4,5-bisphosphate (Li et al., 2015) and extracellular pH (Zhang et al., 2009). In addition, it has been demonstrated that heterologously expressed Kv12.1 and Kv12.2 (and 12.3) α subunits coassemble (Trudeau et al., 1999; Becchetti et al., 2002; Zou et al., 2003; Zhang et al., 2010; Li et al., 2015), although it remains to be determined if heteromeric Kv12.x channels are present in native cells. Interestingly, however, it has been reported that the targeted disruption of the Kcnh3 (Kv12.2) locus increased the input resistances and depolarized the resting membrane potentials of CA1 hippocampal pyramidal neurons (Kazmierczak et al., 2013). In addition, smaller currents were required to evoke action potentials, and the evoked repetitive firing rates were higher in Kv12.2−/− compared with wild-type (WT), CA1 hippocampal neurons (Kazmierczak et al., 2013). The experiments here were designed to test directly the hypothesis that Kv12.1- and/or Kv12.2-encoded K+ channels underlie the subthreshold K+ conductance that reduces the excitability of mouse SCN neurons at night. Combining in vivo molecular genetic strategies to manipulate Kv12.1 or Kv12.2 expression with in vitro electrophysiological and pharmacological approaches to assess the functional consequences of these manipulations, the experiments detailed here demonstrate a critical role for Kv12-encoded K+ channels in regulating the nighttime repetitive firing rates and in controlling the day–night rhythms in the repetitive firing rates of SCN neurons.
Materials and Methods
All reagents were obtained from Sigma-Aldrich unless otherwise noted.
Animals
All procedures involving animals were approved by the Animal Care and Use Committee of Washington University and were conducted in accordance with the United States National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. WT mice were C57BL/6J. The generation of the Kv12.2−/− mouse line, harboring a targeted disruption in the Kcnh3 locus, also maintained in the C57BL/6J background, has been described previously. (Zhang et al., 2010). A similar strategy (illustrated in Fig. S1) was used to generate the Kv12.1−/− line lacking Kcnh8. Crossing Kv12.1−/− and Kv12.2−/− mice provided the Kv12.1−/−/Kv12.2−/− double knockout (DKO) line. All mice were maintained in the C57BL/6J background in one of the Washington University Danforth or Medical School animal facilities.
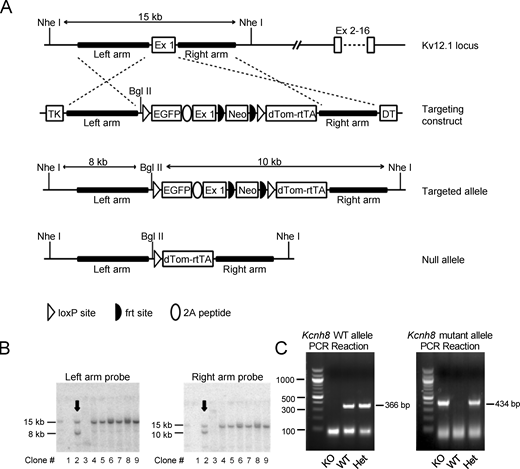
Generation of (Kv12.1−/−) mice harboring a targeted disruption of the Kcnh3 locus. (A) Schematic of the targeted exon 1 Kcnh8 (Ex1) locus, the linearized targeting construct, the initial targeted allele, and the null (knockout) allele generated by Cre-loxP recombination. Targeting of the mouse Kcnh8 (Kv12.1) locus involved homologous recombination (dashed lines) in mouse embryonic stem cells between the native Kcnh8 locus and the targeting vector, and insertion of a loxP site and myristoyl-EGFP into Ex1 immediately upstream of the translation start. A viral 2A sequence joins the myristoyl-EGFP open reading frame to the Kv12.1 open reading frame to potentially allow EGFP-labeling of Kv12.1-expressing cells carrying the targeted allele. Downstream of Ex1 in the first intron, insertions include an frt-bracketed neomycin resistance cassette (Neo) driven by the PGK promoter for positive selection of targeted ES cells on G418, a second loxP site, and dTomato-2A-rtTA (reverse tetracycline trans-activator) cassette (dTOM-rtTA) which includes an SV40 polyadenylation sequence to terminate transcription and block expression of downstream exons. TK (thymidine kinase) and DT (diptheria toxin) expression cassettes flank the left and right, respectively, arms in the targeting construct. These negative selection cassettes are eliminated by homologous recombination but were included to suppress the random insertion of the targeting construct into the ES cell genome. Note the native allele is bracketed by Nhe I restriction sites ∼15 Kb apart. In the targeted allele, the Nhe I sites are preserved, but the distance between them is increased to ∼18 Kb, and a unique Bgl II site is introduced upstream of the 5′ loxP site. Hybridization probes located between either (left or right) arm and the neighboring Nhe I site will label a ∼15 Kb band in a Southern blot of Nhe I/Bgl II digested genomic DNA for the WT allele. The same probes will label bands of 8 Kb (left arm) and 10 Kb (right arm) for the targeted allele. (B) Southern blot analysis of nine G418-resistant ES cell clones following Nhe I/Bgl II digestion of genomic DNA with a probe upstream of the left arm (left) or downstream of the right arm (right). The WT allele is identified by the 15-kb band with both probes, whereas the targeted allele is identified by 8-kb (left arm) and 10-kb (right arm) bands. Arrows indicate the ES cell clone as positive for carrying the targeted allele used to generate the Kv12.1−/− mouse line. Note that DNA isolation failed for two clones (clones #1 and 3). The complete gels from which the “cut outs” in B were derived are provided in the source data. (C) ES clone #2 was karyotyped to confirm chromosome number and morphology and used for injection into C57BL/6J blastocysts. Three male chimeric mice were obtained, two of which transmitted the targeted allele through the germline. Mice carrying the targeted allele were bred with C57BL/6J-TgN(Zp3-Cre)93Knw females, which express Cre-recombinase in the germline (Kemler et al., 2004), to generate heterozygous Kv12.1+/− animals, which were then bred to generate Kv12.1−/− mice. For genotyping, a two-step PCR reaction was used with primers specific to the WT (sense: 5′-TGGTCACAGTGCAGCGGCCAGGGAGTA-3′ and antisense: 5′-AAATTATTGCGCGGATGGAAACAGAGGA-3′) and targeted (sense: 5′-GTCACAGTGCAGCGGCCAGGGAGTAGC-3′ and antisense: 5′-CTTGGCGGTCTGGGTGCCCTCGTAGG-3′) alleles for the Kcnh8 gene. Bands at both 366 (WT) and 434 bp (targeted) identified heterozygous Kv12.1+/− mice; bands at only 366 or 434 bp identified WT or homozygous Kv12.1−/− mice, respectively. Source data are available for this figure: SourceData FS1.
Generation of (Kv12.1−/−) mice harboring a targeted disruption of the Kcnh3 locus. (A) Schematic of the targeted exon 1 Kcnh8 (Ex1) locus, the linearized targeting construct, the initial targeted allele, and the null (knockout) allele generated by Cre-loxP recombination. Targeting of the mouse Kcnh8 (Kv12.1) locus involved homologous recombination (dashed lines) in mouse embryonic stem cells between the native Kcnh8 locus and the targeting vector, and insertion of a loxP site and myristoyl-EGFP into Ex1 immediately upstream of the translation start. A viral 2A sequence joins the myristoyl-EGFP open reading frame to the Kv12.1 open reading frame to potentially allow EGFP-labeling of Kv12.1-expressing cells carrying the targeted allele. Downstream of Ex1 in the first intron, insertions include an frt-bracketed neomycin resistance cassette (Neo) driven by the PGK promoter for positive selection of targeted ES cells on G418, a second loxP site, and dTomato-2A-rtTA (reverse tetracycline trans-activator) cassette (dTOM-rtTA) which includes an SV40 polyadenylation sequence to terminate transcription and block expression of downstream exons. TK (thymidine kinase) and DT (diptheria toxin) expression cassettes flank the left and right, respectively, arms in the targeting construct. These negative selection cassettes are eliminated by homologous recombination but were included to suppress the random insertion of the targeting construct into the ES cell genome. Note the native allele is bracketed by Nhe I restriction sites ∼15 Kb apart. In the targeted allele, the Nhe I sites are preserved, but the distance between them is increased to ∼18 Kb, and a unique Bgl II site is introduced upstream of the 5′ loxP site. Hybridization probes located between either (left or right) arm and the neighboring Nhe I site will label a ∼15 Kb band in a Southern blot of Nhe I/Bgl II digested genomic DNA for the WT allele. The same probes will label bands of 8 Kb (left arm) and 10 Kb (right arm) for the targeted allele. (B) Southern blot analysis of nine G418-resistant ES cell clones following Nhe I/Bgl II digestion of genomic DNA with a probe upstream of the left arm (left) or downstream of the right arm (right). The WT allele is identified by the 15-kb band with both probes, whereas the targeted allele is identified by 8-kb (left arm) and 10-kb (right arm) bands. Arrows indicate the ES cell clone as positive for carrying the targeted allele used to generate the Kv12.1−/− mouse line. Note that DNA isolation failed for two clones (clones #1 and 3). The complete gels from which the “cut outs” in B were derived are provided in the source data. (C) ES clone #2 was karyotyped to confirm chromosome number and morphology and used for injection into C57BL/6J blastocysts. Three male chimeric mice were obtained, two of which transmitted the targeted allele through the germline. Mice carrying the targeted allele were bred with C57BL/6J-TgN(Zp3-Cre)93Knw females, which express Cre-recombinase in the germline (Kemler et al., 2004), to generate heterozygous Kv12.1+/− animals, which were then bred to generate Kv12.1−/− mice. For genotyping, a two-step PCR reaction was used with primers specific to the WT (sense: 5′-TGGTCACAGTGCAGCGGCCAGGGAGTA-3′ and antisense: 5′-AAATTATTGCGCGGATGGAAACAGAGGA-3′) and targeted (sense: 5′-GTCACAGTGCAGCGGCCAGGGAGTAGC-3′ and antisense: 5′-CTTGGCGGTCTGGGTGCCCTCGTAGG-3′) alleles for the Kcnh8 gene. Bands at both 366 (WT) and 434 bp (targeted) identified heterozygous Kv12.1+/− mice; bands at only 366 or 434 bp identified WT or homozygous Kv12.1−/− mice, respectively. Source data are available for this figure: SourceData FS1.
Screening of Kv12.1- and Kv12.2-targeted shRNAs
An interfering RNA strategy (Du et al., 2006) was developed to allow the acute in vivo knockdown of Kv12.1 or Kv12.2 expression selectively in the SCN of adult animals. In initial screening experiments, short hairpin RNA (shRNA) sequences targeting Kv12.1 (n = 5) or Kv12.2 (n = 5), obtained from the RNAi Consortium (TRC) through the Genome Institute at Washington University Medical School, were evaluated in vitro to determine the efficiency of Kv12.1 or Kv12.2 knockdown. For screening, tsA201 cells were cotransfected, using PepMute (Signagen), with a cDNA construct encoding Kv12.1-eYFP or Kv12.2-eYFP and one of the Kv12.1-targeted or Kv12.2-targeted shRNAs. Additional experiments were conducted using a non-targeted (NT), control shRNA generated against a variant of green fluorescent protein. Approximately 48 h following the transfections, cell lysates were prepared, fractionated by SDS-PAGE, transferred to polyvinylidene fluoride (PVDF) membranes, and probed for eYFP expression (#AB3080; polyclonal anti-GFP antibody, 1:1,000; Sigma Aldrich). Following imaging, these blots were probed with an α-tubulin antibody (#AB7291; monoclonal anti-α-tubulin, 1:10,000; Abcam) to verify equal protein loading of each lane and imaged again. The efficiency of the knockdown of Kv12.1-eYFP or Kv12.2-eYFP by each of the Kv12.1- or Kv12.2-targeted shRNAs was quantified by densitometry. The Kv12.1- and Kv12.2-targeted shRNA sequences producing the largest (∼80%) reduction in Kv12.1-eYFP (5′-CAACATTAACTCAGGAGGTTT-3′) or Kv12.2-eYFP (5′-CAGCTTTATGGACCTCCACTT-3′) expression were subsequently evaluated for specificity. In these experiments, tsA201 cells were cotransfected with cDNA constructs encoding Kv12.1-eYFP, Kv12.2-eYFP, or Kv4.1-eYFP, together with the Kv12.1-targeted shRNA, Kv12.2-targeted shRNA, or NT shRNA. 2 d after the transfections, lysates were prepared and fractionated as described above. Western blots were probed first with the polyclonal anti-GFP antibody and imaged, followed by probing with the monoclonal anti-α-tubulin antibody and imaged again.
Generation of shRNA-expressing adeno-associated viruses
The selected Kv12.1-targeted, Kv12.2-targeted, and NT shRNA 21-nucleotide sense sequences were synthesized (Integrated DNA Technologies) into the corresponding 97-nucleotide microRNA-adapted shRNA oligonucleotides containing sense and antisense sequences linked by a 19-nucleotide hairpin loop. Forward and reverse strands were annealed and cloned in a microRNA (human miR30) context into the 3′-untranslated region (3′-UTR) of eGFP in the pPRIME vector (Stegmeier et al., 2005). The entire eGFP-shRNA cassette was then cloned into a viral shuttle vector with a synapsin (SYN) promoter. Adeno-associated viruses, serotype 8 (AAV8), shown previously to produce robust transduction of adult mouse SCN neurons in vivo (Hermanstyne et al., 2017) were generated by the Hope Center Virus Core Facility at Washington University Medical School.
Stereotaxic virus injections
Bilateral stereotaxic injections of the NT, the Kv12.1-targeted, or the Kv12.2-targeted shRNA-expressing AAV8 were made into the SCN of adult (6–10 wk) WT C57BL/6JN male and female mice using previously described methods (Hermanstyne et al., 2017). Briefly, under sterile conditions, each mouse was anesthetized with isoflurane (3%) and secured in a stereotaxic head frame (Kopf Instruments). The head was shaved and Betadine was applied to cleanse and sterilize the shaved region. An incision was then made along the midline and the skin was pulled back to expose the skull. One of the shRNA-expressing viruses was injected (∼600 nl) into each hemisphere of the SCN (coordinates: 0.3 mm rostral to bregma, 0.1 mm left and right to the midline, and 5.6 mm ventral to the pial surface). The injection syringe (Hamilton) delivered the virus at a constant rate of 0.1 μl/min using a syringe pump (KD Scientific). The syringe was left in place for ∼5 min after the injection was completed to minimize the upward reflux of solution during the removal of the needle. Vetabond tissue adhesive (3M) was used to close the incision. Immediately following the surgery, animals were allowed to recover from the anesthesia on a heating pad maintained at 37°C and were given an intraperitoneal (IP) injection of Rimadyl (0.1 ml of 0.05 mg/ml; Pfizer).
Preparation of SCN slices
SCN slices (300 μm) were prepared from adult (8–12 wk old) WT, Kv12.1−/−, Kv12.2−/−, and Kv12.1−/−/Kv12.2−/− (DKO) mice, maintained in either a standard (lights on at 7:00 am and lights off at 7:00 pm) or a reversed (lights on at 7:00 pm and lights off at 7:00 am) 12:12 h light–dark (LD) cycle, using previously described methods (Granados-Fuentes et al., 2012, 2015; Hermanstyne et al., 2017). Zeitgeber times (ZT) are indicated: ZT0 corresponds to the time of lights on and ZT12 to the time of lights off in the animal facility. Daytime slices were routinely prepared at ZT5 from WT, Kv12.1−/−, Kv12.2−/−, and DKO mice maintained in the standard LD cycle, and nighttime slices were routinely prepared at ZT16 from WT, Kv12.1−/−, Kv12.2−/−, and DKO mice maintained in the reversed LD cycle. Using a similar strategy, SCN slices were also prepared from mice 2 wk following NT, Kv12.1-targeted, or Kv12.2-targeted shRNA-expressing AAV8 injections into the SCN. In an additional set of experiments designed to determine if firing rates were affected at any time during the cycle, slices were also obtained from Kv12.1−/− and Kv12.2−/− animals during the transition from “lights-on to lights-off” (ZT12–ZT14), as well as slices prepared mid-day (ZT6–ZT8) and mid-evening (ZT18–ZT20).
For the preparation of daytime slices, brains were rapidly removed (in the light) from animals anesthetized with 1.25% Avertin (Acros Organics, 2,2,2-tribromoethanol and tert-amyl alcohol in 0.9% NaCl; 0.025 ml/g body weight) and placed in ice-cold cutting solution containing (in mM) 240 sucrose, 2.5 KCl, 1.25 NaH2PO4, 25 NaHCO3, 0.5 CaCl2, and 7 MgCl2, saturated with 95% O2/5% CO2. In separate experiments, some daytime slices were prepared at ZT3 and ZT11 to obtain electrophysiological recordings earlier in the day (ZT6–ZT8) and during the transition from lights on to lights off (ZT12–ZT14). For the preparation of nighttime slices, animals in the reversed LD cycle were removed from their cages at ZT15 under infrared illumination, anesthetized with isoflurane, and enucleated using previously described procedure. (Aton et al., 2004; Hattar et al., 2006; Hermanstyne et al., 2016). Following an IP injection of Rimadyl (0.1 ml of 0.05 mg/ml), each animal was allowed to recover from the anesthesia (for ∼1 h) prior to the preparation of slices. At ZT16, animals were anesthetized with 1.25% Avertin; brains were rapidly removed and placed in ice-cold cutting solution. For all experiments, coronal slices (300 µm) were cut on a Leica VT1000 S vibrating blade microtome (Leica Microsystems Inc.) and incubated in a holding chamber with oxygenated artificial cerebrospinal fluid (ACSF) containing (in mM) 125 NaCl, 2.5 KCl, 1.25 NaH2PO4, 25 NaHCO3, 2 CaCl2, 1 MgCl2, and 25 dextrose (∼310 mOsmol l−1), saturated with 95% O2/5% CO2, at room temperature (23–25°C) for at least 1 h before transfer to the recording chamber.
Electrophysiological recordings
Whole-cell current-clamp and action potential- (voltage-) clamp recordings were obtained at room temperature (23–25°C) during the day (ZT7–ZT12) or at night (ZT18–ZT24) from SCN neurons in slices prepared (as described above) during the day or at night from WT, Kv12.1−/−, Kv12.2−/−, or DKO (Kv12.1−/−/Kv12.2−/−) mice. Current-clamp recordings were also obtained from SCN neurons in slices prepared during the day or at night 2 wk following NT, Kv12.1-targeted, or Kv12.2-targeted shRNA-expressing AAV8 injections. In all cases, several animals of each genotype (N = 3–9), with one slice from each animal (N) was used, and electrophysiological data were collected from multiple SCN neurons (n) in each slice. For recordings, SCN neurons were visually identified in slices using differential interference contrast optics with infrared illumination. Slices were perfused continuously with ACSF containing 20 μM Gabazine (Tocris Bioscience) and saturated with 95% O2/5% CO2. Whole-cell current-clamp recordings were obtained using pipettes (4–7 MΩ) containing (in mM) 144 K-gluconate, 10 HEPES, 3 MgCl2, 4 MgATP, 0.2 EGTA, and 0.5 NaGTP (pH 7.3; 300 mOsmol l−1). Using this (K-gluconate) pipette solution, the liquid junction potential was −16.4 mV; junction potentials were not corrected.
For each cell, a loose patch, cell-attached recording was first obtained, and the spontaneous repetitive firing of action potentials was recorded for ∼1 min. This was done routinely prior to establishing the whole-cell recording configuration. Following the formation of a gigaOhm seal, the whole-cell configuration was established, and whole-cell membrane capacitances and series resistances were compensated. Whole-cell spontaneous firing activity was then recorded again for ∼1 min and compared with the data acquired in the cell-attached mode. If any differences in the frequency and/or pattern of repetitive firing in the cell-attached and whole-cell recording configurations were evident, recordings were terminated, and any data acquired from these cells were not included in the analyses. Access resistances were 15–20 MΩ, and data acquisition was terminated if the access resistance increased by ≥20%. Voltage signals were acquired at 100 kHz, filtered at 10 kHz, and stored for offline analysis. Data were collected using a Multiclamp 700B patch clamp amplifier (Molecular Devices) interfaced to a Dell personal computer with a Digidata 1332 and the pCLAMP 10 software package (Molecular Devices). Consistent with previous studies (Itri et al., 2005; Meredith et al., 2006; Kudo et al., 2011; Granados-Fuentes et al., 2015; Hermanstyne et al., 2016; Mazuski et al., 2018), these loose-patch (extracellular) and whole-cell (intracellular) recordings revealed that daytime and nighttime adult mouse SCN neurons display tonic and irregular spontaneous repetitive firing patterns. To facilitate a comparison of repetitive firing rates across groups, the average repetitive firing rate of each cell during the initial 1 min of recording was determined. Mean ± SEM repetitive firing rates for each group were calculated and are reported here. Input resistances (Rin) were determined by measuring the steady-state voltage changes produced by ±5 pA current injections from a membrane potential of −70 mV. The voltage threshold for action potential generation in each cell was determined as the point during the upstroke (depolarizing phase) of the action potential at which the second derivative of the voltage was zero. Resting membrane potentials (Vr) were estimated from phase plots of spontaneous action potentials. For each cell, the first derivative of the membrane potential (dV/dT) was plotted versus the membrane potential (mV), and Vr was defined as the point where dV/dT = 0 during the interspike interval.
For action potential- (voltage-) clamp recordings, 0.1 mM CdCl2 and 1 μM tetrodotoxin (TTX) were added to the standard ACSF bath solution described above. Recording pipettes (3–5 MΩ) contained (in mM) 144 K-gluconate, 10 HEPES, 3 MgCl2, 4 MgATP, 0.2 EGTA, and 0.5 NaGTP (pH 7.3; 300 mOsmol l−1). Spontaneous repetitive firing activity, recorded from a nighttime WT SCN neuron, was used as the voltage command. Tip potentials were zeroed before membrane-pipette seals were formed. Following the formation of a gigaOhm seal and establishing the whole-cell configuration, membrane capacitances and series resistances were compensated electronically. Series resistances were in the range of 15–20 MΩ and were routinely compensated by 70–80%. If the series resistance changed ≥20% during a recording, the experiment was stopped and the acquired data from that cell were not included in the analyses. Outward K+ currents evoked in WT and DKO nighttime and daytime SCN neurons by the action potential voltage command were recorded in TTX- and CdCl2-containing ACSF bath solution before and after application of 20 μM CX4 dissolved in the same (TTX- and CdCl2-containing) ACSF bath solution. The CX4 (1-(2-chloro-6-methylphenyl)-3-(1,2-diphenylethyl) thiourea), developed by Dr. Timothy Jegla and colleagues at the Novartis Research Foundation (La Jolla, CA) and described as a selective blocker of heterologously expressed Kv12.2-encoded K+ currents (Zhang et al., 2010), was provided by Dr. Jegla. Off-line digital subtraction of the outward K+ currents recorded after local application of CX4 from the currents recorded prior to CX4 application provided the CX4-sensitive K+ currents.
Additional experiments were conducted to determine the voltage dependence of activation of the CX4-sensitive currents in nighttime WT SCN neurons. To determine the voltage-dependence of activation of the CX4-sensitive currents, whole-cell Kv currents, evoked in response to 2 s depolarizing voltage steps to potentials between −100 and +50 mV (in 10 mV increments) from a holding potential (HP) of −70 mV, were first recorded in standard ACSF solution containing tetraethylammonium (10 mM), 4-aminopyridine (10 mM), CdCl2 (0.1 mM), and tetrodotoxin (1 μM). Outward Kv currents, evoked using the same protocol, were recorded again (from the same cell) with CX4 (20 μM) added to the same ACSF solution. Off-line digital subtraction of the Kv currents recorded with and without the CX4 in the bath provided the CX4-sensitive currents (ICX4; Fig. S2 A). In each cell, ICX4 conductances at each test potential were calculated and normalized to the maximal conductance (Gmax) determined in the same cell. Mean ± SEM normalized ICX4 conductances (G/Gmax) were then plotted as a function of the test potential and fitted using the Boltzmann equation, G/Gmax = 1 + e[(Va – Vm)/k] (Fig. S2 B), where Va is the voltage of half-maximal activation and k is the slope factor.

CX4-sensitive currents in WT SCN neurons and Cybercyte modeled I Kv12 . (A) Representative whole-cell Kv current recordings obtained from an SCN neuron in an acute slice prepared from a night-phased (ZT19–ZT24) WT SCN are shown. Whole-cell Kv currents, evoked during (2 s) voltage steps to potentials ranging from −100 to +50 mV (in 10 mV increments) from a holding potential of −70 mV, were first recorded in ACSF bath solution with 10 mM TEA and 10 mM 4-AP added (A1) and again following superfusion of the 10 mM TEA- and 10 mM 4-AP-containing ACSF with 20 μM CX4 added (A2). The voltage-clamp paradigm (in gray) is illustrated below the current records. Offline digital subtraction of the records obtained in the presence (A2), from the currents recorded in the absence (A1), of 20 μM CX4 provided the CX4-sensitive currents (A3). ICX4 conductances at each test potential were calculated and normalized to the maximal conductance (Gmax), determined in the same cell. (B) The mean ± SEM normalized conductances of activation of the CX4-sensitive currents are plotted as a function of the test potential and fitted with single Boltzmanns. The V1/2 and the k values derived from these fits for current activation were V1/2 = −4.9 ± 1.0 mV; k = 8.7 ± 1.9 (n = 21). (C) These parameters were used to tune the IK12 model to fit the CX4-sensitive currents that were recorded during action potential–clamp experiments (see Fig. 4). The properties of currents produced by the Cybercyte IKv12 model (lower panel, red) reliably reproduce the CX4-sensitive currents measured in action potential–clamp recordings (middle panel, black) from WT SCN neurons.
CX4-sensitive currents in WT SCN neurons and Cybercyte modeled I Kv12 . (A) Representative whole-cell Kv current recordings obtained from an SCN neuron in an acute slice prepared from a night-phased (ZT19–ZT24) WT SCN are shown. Whole-cell Kv currents, evoked during (2 s) voltage steps to potentials ranging from −100 to +50 mV (in 10 mV increments) from a holding potential of −70 mV, were first recorded in ACSF bath solution with 10 mM TEA and 10 mM 4-AP added (A1) and again following superfusion of the 10 mM TEA- and 10 mM 4-AP-containing ACSF with 20 μM CX4 added (A2). The voltage-clamp paradigm (in gray) is illustrated below the current records. Offline digital subtraction of the records obtained in the presence (A2), from the currents recorded in the absence (A1), of 20 μM CX4 provided the CX4-sensitive currents (A3). ICX4 conductances at each test potential were calculated and normalized to the maximal conductance (Gmax), determined in the same cell. (B) The mean ± SEM normalized conductances of activation of the CX4-sensitive currents are plotted as a function of the test potential and fitted with single Boltzmanns. The V1/2 and the k values derived from these fits for current activation were V1/2 = −4.9 ± 1.0 mV; k = 8.7 ± 1.9 (n = 21). (C) These parameters were used to tune the IK12 model to fit the CX4-sensitive currents that were recorded during action potential–clamp experiments (see Fig. 4). The properties of currents produced by the Cybercyte IKv12 model (lower panel, red) reliably reproduce the CX4-sensitive currents measured in action potential–clamp recordings (middle panel, black) from WT SCN neurons.
Modeling Kv12-encoded (IKv12) and A-type (IA) currents and dynamic clamp recordings
A three-state Markov model of IKv12 channel gating, with two closed states, C1 and C2, and one open state, O, was developed using MATLAB (MathWorks Inc) using published kinetic and steady-state activation and deactivation data for heterologously expressed Kv12.1-encoded channels (Engeland et al., 1998; Scheme 1).

The voltage-dependent forward and reverse transition rates, α and β, between the C2 and O states, determined from the published data (Engeland et al., 1998), were p1exp(p2Vm) and p3exp(−p4Vm), respectively, where Vm is the membrane voltage, p1 = 0.019, p2 = 0.065, p3 = 0.011, and p4 = −0.039 (Clerx et al., 2019). The voltage-independent transition rates (kF and kB) between the C1 and C2 states were approximated from the published time constants of slow activation and the ratio of the fast and slow components of activation of Kv12.1-encoded currents (Engeland et al., 1998). The values determined for kF and kB were 0.008 and 0.0013, respectively. The steady-state voltage-dependence of activation of the model was then adjusted using the voltage-clamp data acquired from SCN neurons with a Kv12-selective inhibitor CX4 (see: Fig. S2, A and B). The Kv12 current is calculated as IKv12 = G*P(O)*(Vm − EK), where G is a scalable parameter for current magnitude, P(O) is the probability that the channel is open, and EK is the reversal potential for K+. Importantly, the currents produced by the Cybercyte IKv12 model reliably reproduce the CX4-sensitive currents measured in action potential–clamp recordings from WT SCN neurons (Fig. S2 C).A Markov model describing the gating of the K+ channels that generate the A-current (IA) was also developed using MATLAB (MathWorks Inc; Scheme 2). The model, which is based on a previously described model of the rapidly activating and inactivating K+ current in (ferret) ventricular myocytes, consists of three closed states, C1–C3, two inactivated states, I0 and I1, and an open (O) state (Campbell et al., 1993), and was populated using acquired voltage-clamp data detailing the time- and voltage-dependent properties of IA in mouse SCN neurons (Hermanstyne et al., 2017).

The voltage-dependent transition rates for IA activation and deactivation, α and β, respectively, determined from analyses of the time constants and sigmoidicity of IA activation in SCN neurons (Hermanstyne et al., 2017), were p1exp(p2Vm) and p3exp(−p2Vm), respectively, where Vm is the membrane voltage, p1 = 0.5, p2 = 0.0328, and p3 = 0.0680. In contrast to activation, the time constants of IA inactivation in mouse SCN neurons were voltage independent over a wide range of membrane potentials (Hermanstyne et al., 2017). This finding suggests a “ball and chain” inactivation mechanism in which a putative “inactivation particle” interacts with the open channel to cause inactivation (Zagotta et al., 1990). In the IA model, this inactivated state is I0, and kF, the voltage-insensitive “on” rate for the particle, is equal to the reciprocal of the inactivation time constant at positive potentials. The unbinding rate of the inactivation particle, kB, was initially estimated by the ability of the model to match steady-state inactivation data for IA (Aldrich et al., 1983). Such a single-state ball and chain model recovers very slowly from inactivation when the membrane is returned to hyperpolarized potentials. To account for the voltage-insensitive inactivation of IA with relatively rapid recovery from inactivation, as observed experimentally (Hermanstyne et al., 2017), we introduced a transition state, I1, into the model. In contrast to I0, the I1 state is a low-affinity binding state in which the energy from the backward movement of the voltage sensor during recovery destabilizes the binding interaction with the inactivation particle in a “push-off ball and chain” inactivation mechanism (Aldrich et al., 1983). The voltage-independent transition rates, kF (0.05), kB (0.00025), kF2 (0.05), and kB2 (0.002), were determined by fitting the data describing steady-state inactivation of IA, and the experimentally determined time constants of IA inactivation and recovery (at −70 mV) from inactivation (Hermanstyne et al., 2017). The ratio factor, F (8), was determined by macroscopic reversibility. The steady-state voltage-dependences of activation and inactivation of the modeled IA are illustrated in Fig. S3 B. The currents are calculated as IA = G*P(O)*(Vm − EK), where G is a scalable parameter for current magnitude, P(O) is the probability that the channel is open, and EK is the reversal potential of K+. As also illustrated in Fig. S3, the simulated IA waveforms evoked in response to depolarizing voltage steps (Fig. S3 C) are similar to IA waveforms recorded from WT SCN neurons (Fig. S3 A).

Cybercyte modeled I A . A Markov model describing the gating of the K+ channels that generate IA in WT SCN neurons was developed based on a previously described model of the rapidly activating and inactivating, IA-like, K+ current in (ferret) ventricular myocytes (Campbell et al., 1993), and was populated using previously acquired voltage-clamp data detailing the time- and voltage-dependent properties of IA in mouse SCN neurons (Hermanstyne et al., 2017). (A) Representative IA waveforms recorded from a WT SCN neuron (Hermanstyne et al., 2017) in response to voltage steps to test potentials ranging from −40 to +40 mV (in 5 mV increments) from a HP of −70 mV are shown; the voltage-clamp paradigm is illustrated above the current records. (B) The voltage-dependences of activation and inactivation for IA in WT SCN neurons (Hermanstyne et al., 2017) were determined using protocols identical to those described above for the CX4-senstive currents. The V1/2 and the k values derived from these fits for current activation (open symbols) and inactivation (closed symbols) were V1/2 = −9.3 ± 1.3 mV; k = 12.7 ± 0.8 (n = 12) and V1/2 = −59.4 ± 2.1 mV, k = 7.9 ± 0.7 (n = 12), respectively. (C) These parameters were used to tune the model. (C) The waveforms of the currents produced by the Cybercyte IA model reliably reproduced IA recorded from WT SCN neurons (A).
Cybercyte modeled I A . A Markov model describing the gating of the K+ channels that generate IA in WT SCN neurons was developed based on a previously described model of the rapidly activating and inactivating, IA-like, K+ current in (ferret) ventricular myocytes (Campbell et al., 1993), and was populated using previously acquired voltage-clamp data detailing the time- and voltage-dependent properties of IA in mouse SCN neurons (Hermanstyne et al., 2017). (A) Representative IA waveforms recorded from a WT SCN neuron (Hermanstyne et al., 2017) in response to voltage steps to test potentials ranging from −40 to +40 mV (in 5 mV increments) from a HP of −70 mV are shown; the voltage-clamp paradigm is illustrated above the current records. (B) The voltage-dependences of activation and inactivation for IA in WT SCN neurons (Hermanstyne et al., 2017) were determined using protocols identical to those described above for the CX4-senstive currents. The V1/2 and the k values derived from these fits for current activation (open symbols) and inactivation (closed symbols) were V1/2 = −9.3 ± 1.3 mV; k = 12.7 ± 0.8 (n = 12) and V1/2 = −59.4 ± 2.1 mV, k = 7.9 ± 0.7 (n = 12), respectively. (C) These parameters were used to tune the model. (C) The waveforms of the currents produced by the Cybercyte IA model reliably reproduced IA recorded from WT SCN neurons (A).
A dynamic clamp was carried out using a commercially available Cybercyte Dynamic Clamp System, Cybercyte DC1 (Bett et al., 2013; Du et al., 2021) from Cytocybernetics. This system consists of a 16-channel, 16-bit, 100 kS/s MCC PCIe-DAS1602/16 board installed and configured in a Dell 5820 Precision workstation. The average loop time determined for this system was 22 μs. Prior to experiments on SCN neurons, the IK12 and IA formulations were converted to Cybercyte channel definition files using Cybersolver software and validated by applying the simulated currents to a model cell (Molecular Devices) with an input resistance of 500 MΩ and a capacitance of 30 pF in whole-cell mode. The Cybercyte DC1 system allows current amplitudes to be scaled during dynamic clamp experiments such that the modeled currents (IK12 and IA) can be “added” or “subtracted” (by the addition of IK12 or IA of the opposite polarity) in real-time during whole-cell current-clamp recordings from nighttime and daytime WT SCN neurons.
Initial experiments were undertaken on nighttime WT SCN neurons to determine the minimal subtracted IKv12 amplitude that increased the spontaneous repetitive firing rates of most (9 of 10) cells. These experiments revealed that the minimal subtracted (peak) IKv12 amplitude required to affect repetitive firing rates in most (9 of 10) nighttime WT SCN neurons was 2 pA, and this value was set as x. The effects of subtracting varying amplitudes (in integral multiples of x) of IKv12 on the repetitive firing rates of WT nighttime neurons were then determined. For daytime WT neurons, IKv12 was scaled (to 0.5 pA) to reflect the day–night difference in the (mean) CX4-sensitive current amplitudes measured in WT neurons (Fig. 4 D), and the effects of the subtraction of integral multiples of this value (i.e., x = 0.5 pA) on the repetitive firing rates of daytime WT neurons were determined. A similar strategy was used in dynamic clamp experiments to determine the effects of adding IKv12 in nighttime and daytime WT neurons, as well as of subtracting/adding IA in nighttime and daytime WT SCN neurons. Preliminary experiments revealed that the minimal added IKv12 amplitude required to affect repetitive firing rates in most (9 of 10) nighttime WT SCN neurons was 5 pA and IKv12 was scaled (to 2 pA), again to reflect the day–night difference in CX4-sensitive current amplitudes measured in WT neurons (Fig. 4 D). The minimal modeled IA amplitude required to alter repetitive firing rates in most (9 of 10) nighttime WT SCN neurons was 20 pA. In addition, because previous studies demonstrated that IA densities in WT nighttime and daytime SCN neurons are not significantly different (Hermanstyne et al., 2017), no day–night scaling factor was used.
For dynamic clamp recordings, the membrane voltage was sampled at 20 kHz and the Cybercyte DC1 returned the corresponding modeled IKv12 (or IA) at a rate of 50 kHz. For each cell, spontaneous repetitive firing was recorded for 1 min. Modeled currents were then added or subtracted in multiples (i.e., x, 2x, −x, −2x, etc.) of the minimal (x) amplitudes, determined as described above, and the resulting repetitive firing rates were measured. The percent change in the repetitive firing rate with each current injection was then determined in each cell. These values were then averaged across cells and the mean ± SEM percent changes in firing rates as a function of the injected current amplitudes are presented.
Analyses of wheel-running activity
Adult (9–12 wk old) WT, Kv12.1−/−, Kv12.2−/−, and DKO mice were placed (individually) in cages equipped with running wheels in light-tight chambers illuminated with fluorescent bulbs (2.4 ± 0.5 × 1018 photons/s*m2; General Electric). Wheel-running activity was recorded (Clocklab Actimetrics) in 6-min bins for 10 d in a 12:12 h LD cycle, followed by recordings in constant darkness (DD) for at least 20 d. Similar recordings were obtained from animals 10 d following bilateral injections (600 nl in each hemisphere) of the NT, Kv12.1-targeted, or Kv12.2-targeted shRNA-expressing AAV8 into the SCN of adult (9–12 wk) WT mice. The period of rhythmicity of each mouse was determined using χ2 periodogram analysis (Sokolove and Bushell, 1978) of continuous recordings for 10 d in DD (Clocklab Actimetrics). Wheel-running was considered rhythmic if the χ2 periodogram value exceeded the 99.9% confidence interval (Qp value). Statistical analysis of the circadian periods was compared between groups using a one-way analysis of variance (ANOVA).
Quantitative RT-PCR analysis
Total RNA was isolated from (300 μm) SCN slices, collected every 4 h (ZT 3, 7, 11, 15, 19, and 23) for two consecutive days from adult (8–10 wk) WT mice (n = 7–8), maintained in standard and reversed LD conditions, and DNase treated using previously described methods (Marionneau et al., 2008). RNA concentrations were determined by optical density measurements (Marionneau et al., 2008). The mRNA transcript expression levels of genes encoding Per2, Bmal, Kcnh8 (Kv12.1), Kcnh3 (Kv12.2), as well as of the endogenous control gene Hprt (hypoxanthine guanine phosphoribosyl transferase), were determined using Taqman-based real-time quantitative (RT) PCR. Data were collected with instrument spectral compensations using the Applied Biosystems SDS 2.2.2 software and analyzed using the threshold cycle (CT) relative quantification method (Schmittgen and Livak, 2008). The expression of each transcript was normalized to the expression of Hprt in the same sample and evaluated for rhythmicity using JTK cycle analysis (Hughes et al., 2010) with the period set to 24 h. The primers sequences used were Per2: 5′-TCCACCGGCTACTGATGCA-3′ and 5′-TGGATGATGTCTGGCTCATGA-3′; Bmal: 5′-GTAGGATGTGACCGAGGGAAGA-3′ and 5′-AGTCAAACAAGCTCTGGCCAAT-3′; Kcnh8 (Kv12.1): 5′-AGGATTACTGGCGCCACAGA-3′ and 5′-CTTTGCCACTTGGGCATTG-3′; Kcnh3 (Kv12.2): 5′-GCAACGTGTCCGCTAACACA-3′ and 5′-GCCGTCACATTCCCAAACA-3′; and Hprt: 5′-TGAATCACGTTTGTGTCATTAGTGA-3′ and 5′-TTCAACTTGCGCTCATCTTAGG-3′.
Statistical analysis
Electrophysiological data were compiled and analyzed using ClampFit (v. 10.2; Molecular Devices), Mini Analysis (v. 6.0.7; Synaptosoft), and Prism (v. 8.2; GraphPad Software). Averaged data are presented as means ± standard error of the means (SEM). Statistical analyses were performed using Student’s t test or one-way ANOVA with Newman-Kuels post-hoc pairwise comparisons, as indicated in the text, figure legends, or tables; P values are reported.
Online supplemental material
Fig. S1 illustrates the strategy used to generate Kv12.1−/− mouse line, together with the validation of this mouse line. Fig. S2 shows the waveforms of the CX4-sensitive K+ currents in WT SCN neurons and the Cybercyte modeled Kv12 currents, IKv12, and Fig. S3 shows the Cybercyte modeled IA. In Fig. S4, the average repetitive firing rates and membrane properties determined in individual SCN slices from WT, Kv12.1−/−, Kv12.2−/−, NT shRNA-expressing, Kv12.1 shRNA-expressing, and Kv12.2 shRNA expressing animals are shown. The repetitive firing rates determined in WT, Kv12.1−/−, and Kv12.2−/− SCN neurons throughout the day and night are presented in Fig. S5. The effects of the dynamic clamp-mediated addition of IKv12 on the repetitive firing rates of nighttime WT and Kv12.1−/− SCN neurons are compared in Fig. S6. The kinetic differences between the modeled IA and IKv12 are illustrated in Fig. S7 and, in Fig. S8, the effects of dynamic clamp-mediated subtraction of IA and IKv12 versus IA are compared directly. In Fig. S9, the variations in the transcript expression levels of Kcnh8 (Kv12.1), Kcnh3 (Kv12.2), as well as of Per and Bmal, measured over a 48-h period at 4-h intervals, are presented. Table S1 presents the data represented in Fig. S4.
Results
Targeted deletion of Kcnh8 or Kcnh3 selectively alters nighttime firing rates in SCN neurons
To explore the roles of Kv12.1/Kv12.2-encoded K+ channels in regulating spontaneous repetitive firing in the SCN, we obtained whole-cell current-clamp recordings during the day (ZT7–ZT12) and at night (ZT19–ZT24) from SCN neurons in slices prepared from adult WT mice and from animals harboring targeted disruptions in the Kcnh8 (Kv12.1−/−) or the Kcnh3 (Kv12.2−/−) locus (Fig. 1). Representative recordings from daytime (Fig. 1 A) and nighttime (Fig. 1 B) WT, Kv12.1−/−, and Kv12.2−/− SCN neurons are shown in Fig. 1. Consistent with previous reports (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020), these experiments revealed a large (P = 0.0001, one-way ANOVA) day–night difference in the mean ± SEM repetitive firing rates of WT SCN neurons (Fig. 1 C and Table 1). In marked contrast, there were no measurable differences in the mean ± SEM daytime and nighttime repetitive firing rates of Kv12.1−/− (P = 0.99, one-way ANOVA) or Kv12.2−/− (P = 0.79, one-way ANOVA) SCN neurons (Fig. 1 C and Table 1). In addition, the loss of the day–night difference in the mean repetitive firing rates of Kv12.1−/− and Kv12.2−/− SCN neurons reflects a selective increase (compared with WT cells) in mean nighttime repetitive firing rates (Fig. 1 C and Table 1); the daytime repetitive firing rates of Kv12.1−/− and Kv12.2−/− SCN neurons are similar to WT cells (Fig. 1 C and Table 1). Similar results were obtained in recordings from SCN neurons in slices prepared from Kv12.1−/−/Kv12.2−/− (DKO) animals lacking both Kv12.1 and Kv12.2.

The repetitive firing rates of Kv12.1 −/− and Kv12.2 −/− SCN neurons are similar during the day and at night. (A and B) Representative daytime (A) and nighttime (B) whole-cell current-clamp recordings from WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) SCN neurons are shown. (C) Daytime and nighttime repetitive firing rates, measured in individual WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) SCN neurons are plotted; mean ± SEM repetitive firing rates are indicated; n = numbers of cells. As reported previously (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020), the mean ± SEM repetitive firing rate in WT SCN neurons is higher (P = 0.0001, one-way ANOVA) during the day than at night (Table 1). In contrast, there are no day–night differences (one-way ANOVA) in the mean ± SEM repetitive firing rates of Kv12.1−/− (red squares; P = 0.99) or Kv12.2−/− (blue squares; P = 0.79) SCN neurons (Table 1). (D and E) Input resistances (D) and membrane potentials (E), measured in individual WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) SCN neurons during the day and at night, are plotted; mean ± SEM values and P values are indicated. In contrast to WT SCN neurons, there are no day–night differences (one-way ANOVA) in the mean ± SEM input resistances (D) or membrane potentials (E) of Kv12.1−/− (red squares) or Kv12.2−/− (blue squares) SCN neurons (Table 1).
The repetitive firing rates of Kv12.1 −/− and Kv12.2 −/− SCN neurons are similar during the day and at night. (A and B) Representative daytime (A) and nighttime (B) whole-cell current-clamp recordings from WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) SCN neurons are shown. (C) Daytime and nighttime repetitive firing rates, measured in individual WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) SCN neurons are plotted; mean ± SEM repetitive firing rates are indicated; n = numbers of cells. As reported previously (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020), the mean ± SEM repetitive firing rate in WT SCN neurons is higher (P = 0.0001, one-way ANOVA) during the day than at night (Table 1). In contrast, there are no day–night differences (one-way ANOVA) in the mean ± SEM repetitive firing rates of Kv12.1−/− (red squares; P = 0.99) or Kv12.2−/− (blue squares; P = 0.79) SCN neurons (Table 1). (D and E) Input resistances (D) and membrane potentials (E), measured in individual WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) SCN neurons during the day and at night, are plotted; mean ± SEM values and P values are indicated. In contrast to WT SCN neurons, there are no day–night differences (one-way ANOVA) in the mean ± SEM input resistances (D) or membrane potentials (E) of Kv12.1−/− (red squares) or Kv12.2−/− (blue squares) SCN neurons (Table 1).
Resting and active membrane properties of WT, Kv12.1−/−, Kv12.2−/−, and DKO (Kv12.1−/−/Kv12.2−/−) SCN neurons during the day and at night
| Firing rate (Hz) | Rin (GΩ) | Vr (mV) | ISI (ms) | ||
|---|---|---|---|---|---|
| Day ZT 7–12 | WT | 4.0 ± 0.5 n = 28 | 1.7 ± 0.2** n = 20 | −41.6 ± 0.7*** n = 28 | 280 ± 9* n = 28 |
| Kv12.1−/− | 4.3 ± 0.6 n = 38 | 2.0 ± 0.2 n = 33 | −40.4 ± 0.7 n = 38 | 252 ± 9 n = 38 | |
| Kv12.2−/− | 3.6 ± 0.3 n = 35 | 2.0 ± 0.1 n = 34 | −38.4 ± 0.7 n = 35 | 311 ± 9 n = 35 | |
| DKO | 4.1 ± 0.3 n = 6 | 1.6 ± 0.2 n = 6 | −40.8 ± 1.6 n = 6 | 279 ± 13 n = 6 | |
| Night ZT 18–24 | WT | 0.8 ± 0.2* n = 25 | 1.0 ± 0.1** n = 24 | −46.7 ± 1.0*** n = 25 | 418 ± 29* n = 20 |
| Kv12.1−/− | 4.0 ± 0.7 n = 21 | 1.8 ± 0.2 n = 18 | −37.7 ± 1.2 n = 21 | 254 ± 6 n = 21 | |
| Kv12.2−/− | 3.1 ± 0.5 n = 22 | 2.0 ± 0.2 n = 22 | −37.5 ± 0.9 n = 22 | 319 ± 17 n = 22 | |
| DKO | 3.4 ± 0.4 n = 6 | 1.5 ± 0.2 n = 6 | −40.5 ± 3.2 n = 6 | 318 ± 12 n = 6 | |
Values are means ± SEM; n = number of cells; Rin = input resistance; Vr = resting membrane potentials; ISI = interspike interval. Values in daytime and nighttime WT neurons are significantly different (*P = 0.0001; **P = 0.02; ***P = 0.0005).
As is evident in Fig. 1 C, the repetitive firing rates of WT, as well as Kv12.1−/− and Kv12.2−/−, SCN neurons are quite variable. Similar variability in (in daytime and nighttime) repetitive firing rates for all genotypes was evident across slices/animals. Importantly, comparisons of the averaged data acquired in individual animals (N; Fig. S4) also revealed a large (P = 0.0001, one-way ANOVA) day–night difference in the mean repetitive firing rates in WT SCN slices (Fig. S4 A and Table S1), whereas there were no day–night differences in mean repetitive firing rates determined in SCN slices prepared from Kv12.1−/− (P = 0.84, one-way ANOVA) or Kv12.2−/− (P = 0.55, one-way ANOVA) animals (Fig. 1 C, Table 1, Fig. S4, and Table S1).
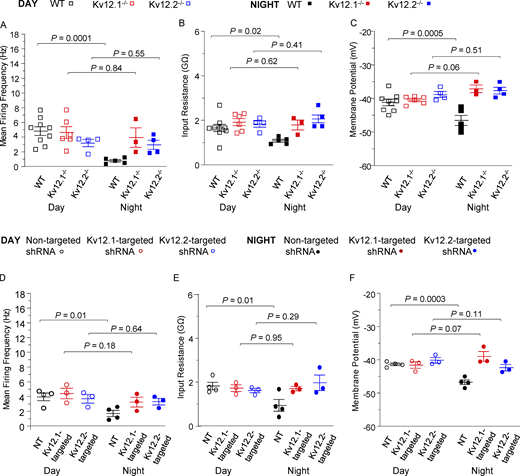
Repetitive firing rates in individual Kv12.1−/−, Kv12.2−/−, Kv12.1-targeted shRNA-expressing, and Kv12.2-targeted shRNA-expressing animals/slices. Comparisons of repetitive firing rates and membrane properties of WT (black squares), Kv12.1−/− (red squares), Kv12.2−/− (blue squares), NT shRNA-expressing (black circles), Kv12.1-targeted shRNA-expressing (red circles), and Kv12.2-targeted shRNA-expressing (blue circles) daytime (ZT 7–ZT 12; open symbols) and nighttime (ZT 18–ZT 24; closed symbols) SCN neurons; the individual data points represent the average values determined in slices from individual animals; N = number of animals. (A–C) Repetitive firing rates (A), input resistances (B), and membrane potentials (C) were measured in WT, Kv12.1−/−, and Kv12.2−/− SCN neurons in slices prepared during the day (open symbols: N = 9 for WT [black square]; N = 6 for Kv12.1−/− [red square]; N = 4 for Kv12.2−/− [blue square]) or at night (closed symbols: N = 5 for WT [black square]; N = 3 for Kv12.1−/− [red square]; N = 4 for Kv12.2−/− [blue square]) and average values (in individual slices/animals) are plotted. (D–F) Average daytime and nighttime repetitive firing rates (D), input resistances (E), and membrane potentials (F), determined in daytime (open symbols) NT shRNA- (black circles; N = 4), Kv12.1-targeted shRNA- (red circles; N = 3), Kv12.2-targeted shRNA- (blue circles; N = 3), and nighttime (closed symbols) NT shRNA- (black circles; N = 4), Kv12.1-targeted shRNA- (red circles; N = 3), and Kv12.2-targeted shRNA- (blue circles; N = 3) expressing SCN neurons are presented. In A–F, mean ± SEM values are plotted and P values are indicated. All data are also tabulated in Table S1.
Repetitive firing rates in individual Kv12.1−/−, Kv12.2−/−, Kv12.1-targeted shRNA-expressing, and Kv12.2-targeted shRNA-expressing animals/slices. Comparisons of repetitive firing rates and membrane properties of WT (black squares), Kv12.1−/− (red squares), Kv12.2−/− (blue squares), NT shRNA-expressing (black circles), Kv12.1-targeted shRNA-expressing (red circles), and Kv12.2-targeted shRNA-expressing (blue circles) daytime (ZT 7–ZT 12; open symbols) and nighttime (ZT 18–ZT 24; closed symbols) SCN neurons; the individual data points represent the average values determined in slices from individual animals; N = number of animals. (A–C) Repetitive firing rates (A), input resistances (B), and membrane potentials (C) were measured in WT, Kv12.1−/−, and Kv12.2−/− SCN neurons in slices prepared during the day (open symbols: N = 9 for WT [black square]; N = 6 for Kv12.1−/− [red square]; N = 4 for Kv12.2−/− [blue square]) or at night (closed symbols: N = 5 for WT [black square]; N = 3 for Kv12.1−/− [red square]; N = 4 for Kv12.2−/− [blue square]) and average values (in individual slices/animals) are plotted. (D–F) Average daytime and nighttime repetitive firing rates (D), input resistances (E), and membrane potentials (F), determined in daytime (open symbols) NT shRNA- (black circles; N = 4), Kv12.1-targeted shRNA- (red circles; N = 3), Kv12.2-targeted shRNA- (blue circles; N = 3), and nighttime (closed symbols) NT shRNA- (black circles; N = 4), Kv12.1-targeted shRNA- (red circles; N = 3), and Kv12.2-targeted shRNA- (blue circles; N = 3) expressing SCN neurons are presented. In A–F, mean ± SEM values are plotted and P values are indicated. All data are also tabulated in Table S1.
Also consistent with previous reports (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020), there were marked day–night differences in the mean ± SEM input resistances (Fig. 1 D; P = 0.02, one-way ANOVA), resting membrane potentials (Fig. 1 E; P = 0.0005, one-way ANOVA), and interspike intervals (Table 1; P = 0.001, one-way ANOVA) of WT SCN neurons (Table 1). In contrast, the mean ± SEM input resistances, resting membrane potentials, and interspike intervals measured in daytime and nighttime Kv12.1−/−, Kv12.2−/−, or DKO SCN neurons were not significantly different (Fig. 1, D and E; and Table 1). Similar results were obtained when the average data acquired in SCN slices from individual animals were compared (Fig. S4 and Table S1). Consistent with the repetitive firing data, these results reflect the higher mean input resistances and depolarized membrane potentials of nighttime Kv12.1−/−, Kv12.2−/−, and DKO compared with nighttime WT, SCN neurons (Fig. 1, D and E; Table 1; and Table S1). The mean ± SEM daytime input resistances and resting membrane potentials of Kv12.1−/−, Kv12.2−/−, and DKO SCN neurons are similar to those measured in daytime WT SCN neurons (Fig. 1, D and E; and Table 1).
Because the majority of the data presented in Fig. 1 and Table 1 was obtained during the latter portion of the night (i.e., ZT20–ZT24), additional recordings were obtained throughout the light–dark cycle. Specifically, additional cell-attached current-clamp recordings were obtained from cells in slices during the transition from lights-on to lights-off (ZT12–ZT14), as well as slices prepared mid-day (ZT6–ZT8) and mid-evening (ZT18–ZT20). Analysis of these recordings revealed that during the transition from day to night, the mean repetitive firing rate of WT SCN neurons decreased to ∼1 Hz (compared to ∼4 Hz at mid-day), and this rate was maintained to the end of the night phase (Fig. S5). In marked contrast, the mean repetitive firing rates of Kv12.1−/− and Kv12.2−/− SCN neurons during the “day to night” transition (∼3.5 Hz) were much higher (Fig. S5) and remained high at midday and mid-evening (Fig. S5). In addition, the mean ± SEM daytime repetitive firing rates of Kv12.1−/− and Kv12.2−/− SCN neurons are indistinguishable from WT SCN cells (Fig. 1 C and Table 1). Similar results were obtained in recordings from DKO SCN neurons (Table 1). Consistent with the similarities in repetitive firing rates measured during the day and at night, Kv12.1−/−, Kv12.2−/−, and DKO SCN neurons do not display the day–night difference in interspike intervals characteristic of WT SCN neurons (Table 1). Taken together, these results demonstrate that the repetitive firing rates of Kv12.1−/− and Kv12.2−/− SCN neurons are increased throughout the night.

Repetitive firing rates of Kv12.1 −/− and Kv12.2 −/− SCN neurons are higher than in WT SCN neurons throughout the night. Cell-attached voltage recordings were obtained for WT, Kv12.1−/−, and Kv12.2−/− SCN neurons in acute slices prepared at various times throughout the circadian cycle. As is evident, mean ± SEM peak firing rates were high throughout the day (yellow shaded region) in WT (black circles) SCN neurons (n = 132), subsequently decreased during the transition from day to night, and remained low throughout the night. Conversely, the mean ± SEM peak firing rates of Kv12.1−/− (red circles; n = 121) and Kv12.2−/− (blue circles; n = 125) SCN neurons did not vary measurably over time and were consistently high (similar to the daytime repetitive firing rates of WT SCN neurons) throughout the day and night.
Repetitive firing rates of Kv12.1 −/− and Kv12.2 −/− SCN neurons are higher than in WT SCN neurons throughout the night. Cell-attached voltage recordings were obtained for WT, Kv12.1−/−, and Kv12.2−/− SCN neurons in acute slices prepared at various times throughout the circadian cycle. As is evident, mean ± SEM peak firing rates were high throughout the day (yellow shaded region) in WT (black circles) SCN neurons (n = 132), subsequently decreased during the transition from day to night, and remained low throughout the night. Conversely, the mean ± SEM peak firing rates of Kv12.1−/− (red circles; n = 121) and Kv12.2−/− (blue circles; n = 125) SCN neurons did not vary measurably over time and were consistently high (similar to the daytime repetitive firing rates of WT SCN neurons) throughout the day and night.
Acute in vivo knockdown of Kv12.1 or Kv12.2 also selectively increases nighttime firing rates
The electrophysiological studies above demonstrate increased repetitive firing rates and input resistances, reduced interspike intervals, and depolarized resting membrane potentials in nighttime Kv12.1−/−, Kv12.2−/−, and DKO, compared with WT, SCN neurons; the repetitive firing and intrinsic membrane properties of daytime Kv12.1−/−, Kv12.2−/−, DKO, and WT SCN neurons, in contrast, are indistinguishable (Fig. 1, Fig. S4, Table 1, and Table S1). Interpreting the physiological significance and the functional implications of these findings is confounded, however, by the fact that the Kv12.1−/−, Kv12.2−/−, and DKO mice lack Kv12.1 or/and Kv12.2 throughout development. Additional experiments were undertaken, therefore, to test the hypotheses that Kv12.1 and/or Kv12.2 are required for the regulation of the nighttime repetitive firing rates and the membrane properties of mature SCN neurons using an interfering RNA (Du et al., 2006) strategy to knockdown the expression of the Kv12.1 and Kv12.2 subunits acutely in the SCN of adult animals in vivo.
To identify shRNA sequences that effectively reduce Kv12.1 (or Kv12.2) expression, five Kv12.1-targeted (or five Kv12.2-targeted) shRNAs were screened individually in tsA-201 cells cotransfected with a cDNA construct encoding Kv12.1-eYFP (or Kv12.2-eYFP). The Kv12.1-targeted (and Kv12.2-targeted shRNA) sequences providing the maximal (∼80%) reductions in Kv12.1 (or Kv12.2) expression identified in these initial screens (see Materials and methods) were further evaluated for specificity. In these experiments, tsA-201 cells were cotransfected with the validated Kv12.1-targeted or Kv12.2-targeted shRNA or a NT shRNA with cDNA constructs encoding Kv12.1-eYFP, Kv12.2-eYFP, or Kv4.1-eYFP (from another Kv subfamily). As illustrated in Fig. 2, Western blot analysis revealed that the Kv12.1-targeted shRNA markedly reduced the expression of Kv12.1-eYFP (Fig. 2 A), without measurably affecting Kv12.2-eYFP or Kv4.1-eYFP (Fig. 2, B and C). Similarly, the Kv12.2-targeted shRNA markedly reduced Kv12.2-eYFP, without measurably affecting Kv12.1-eYFP or Kv4.1-eYFP (Fig. 2, A–C). These selective Kv12.1-targeted and Kv12.2-targeted shRNAs, as well as the NT shRNA, were then cloned (separately), in a microRNA (human miR30) context (Stegmeier et al., 2005), into a plasmid containing a synapsin promoter and eGFP (Hermanstyne et al., 2017), and adeno-associated viruses serotype 8 (AAV8) were generated.
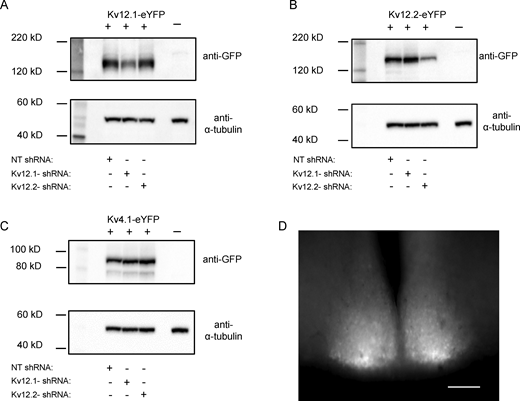
Validation of the Kv12.1-targeted and Kv12.2-targeted shRNAs. To examine the specificity of the selected Kv12.1-targeted and Kv12.2-targeted shRNAs, each was expressed in tsA-201 cells with a cDNA construct encoding Kv12.1-eYFP, Kv12.2-eYFP, or Kv4.1-eYFP; parallel experiments were completed with the NT shRNA. Approximately 24 h following transfections, cell lysates were prepared, fractionated by SDS-PAGE, transferred to PVDF membranes, and probed with an anti-GFP antibody. All blots were also subsequently probed with an anti-α-tubulin antibody to verify equal protein loading. (A) Co-expression with the Kv12.1-targeted shRNA markedly reduced Kv12.1-eYFP protein expression, whereas the Kv12.2-targeted shRNA and the NT shRNA were without effects on Kv12.1-eYFP. (B) Similarly, co-expression with the Kv12.2-targeted shRNA, but not the Kv12.1-targeted or the NT shRNA, reduced Kv12.2-eYFP protein expression. (C) In contrast, neither the Kv12.1-targeted nor the Kv12.2-targeted shRNA measurably affected expression of Kv4.1-eYFP. The complete blots from which the “cut outs” shown in A–C were derived are provided in the source data. (D) eGFP-expressing neurons were readily identified in acute SCN slices prepared 2 wk following bilateral injections of the Kv12.1-targeted (or the Kv12.2-targeted) shRNA-expressing AAV8 into the SCN; scale bar = 250 μm. Source data are available for this figure: SourceData F2.
Validation of the Kv12.1-targeted and Kv12.2-targeted shRNAs. To examine the specificity of the selected Kv12.1-targeted and Kv12.2-targeted shRNAs, each was expressed in tsA-201 cells with a cDNA construct encoding Kv12.1-eYFP, Kv12.2-eYFP, or Kv4.1-eYFP; parallel experiments were completed with the NT shRNA. Approximately 24 h following transfections, cell lysates were prepared, fractionated by SDS-PAGE, transferred to PVDF membranes, and probed with an anti-GFP antibody. All blots were also subsequently probed with an anti-α-tubulin antibody to verify equal protein loading. (A) Co-expression with the Kv12.1-targeted shRNA markedly reduced Kv12.1-eYFP protein expression, whereas the Kv12.2-targeted shRNA and the NT shRNA were without effects on Kv12.1-eYFP. (B) Similarly, co-expression with the Kv12.2-targeted shRNA, but not the Kv12.1-targeted or the NT shRNA, reduced Kv12.2-eYFP protein expression. (C) In contrast, neither the Kv12.1-targeted nor the Kv12.2-targeted shRNA measurably affected expression of Kv4.1-eYFP. The complete blots from which the “cut outs” shown in A–C were derived are provided in the source data. (D) eGFP-expressing neurons were readily identified in acute SCN slices prepared 2 wk following bilateral injections of the Kv12.1-targeted (or the Kv12.2-targeted) shRNA-expressing AAV8 into the SCN; scale bar = 250 μm. Source data are available for this figure: SourceData F2.
2 wk following stereotaxic injections of the Kv12.1-targeted, Kv12.2-targeted, or NT shRNA-expressing AAV8 into the adult SCN, whole-cell current-clamp recordings were obtained from eGFP-positive SCN neurons in slices (Fig. 2 D). Representative recordings from NT, Kv12.1-targeted, and Kv12.2-targeted shRNA-expressing SCN neurons during the daytime (ZT7–ZT12) and nighttime (ZT18–ZT24) are presented in Fig. 3, A and B. Analyses of the repetitive firing data acquired from many daytime and nighttime eGFP-positive SCN neurons revealed a day–night difference in the mean ± SEM repetitive firing rates of NT shRNA-expressing SCN neurons (Fig. 3 C and Table 2). Similar to WT neurons (Table 1), the mean ± SEM repetitive firing rate measured in NT shRNA-expressing SCN neurons was much (P = 0.0001, one-way ANOVA) higher during the day than at night (Fig. 3 C and Table 2). In contrast, the mean ± SEM repetitive firing rates of both Kv12.1-targeted shRNA- (P = 0.11) and Kv12.2-targeted (P = 0.84) shRNA-expressing SCN neurons were similar day and night. The lack of day–night differences in the mean ± SEM repetitive firing rates of Kv12.1-targeted and Kv12.2-targeted shRNA-expressing SCN neurons reflects the fact that the nighttime firing rates in these cells were high compared with nighttime NT shRNA-expressing SCN neurons (Fig. 3 C and Table 2).

Acute in vivo shRNA-mediated knockdown of Kv12.1 or Kv12.2 increases the nighttime, but not the daytime, repetitive firing rates of SCN neurons. (A and B) Representative daytime (A) and nighttime (B) whole-cell current-clamp recordings obtained from NT shRNA- (black circles), Kv12.1-targeted shRNA- (red circles), and Kv12.2-targeted shRNA- (blue circles) expressing SCN neurons during the day (ZT7–ZT12) and at night (ZT18–ZT24) are shown. (C) Daytime and nighttime repetitive firing rates measured in individual SCN neurons expressing the NT (black circles), Kv12.1-targeted (red circles), or Kv12.2-targeted shRNA (blue circles) are plotted; mean ± SEM repetitive firing rates are indicated. Similar to WT SCN[7–10] neurons (Fig. 1), the mean ± SEM repetitive firing rate of NT shRNA-expressing (black circles) SCN neurons was lower (P = 0.0001, one-way ANOVA) at night than during the day (Table 2). In marked contrast, the mean ± SEM repetitive firing rates of Kv12.1- (red circles) and Kv12.2- (blue circles) targeted shRNA-expressing SCN neurons at night are not (one-way ANOVA) different from daytime firing rates (Table 2). (D and E) The input resistances (D) and membrane potentials (E) of NT shRNA- (black circles), Kv12.1-targeted shRNA- (red circles), and Kv12.2-targeted shRNA-expressing (blue circles) SCN neurons during the day and at night were also determined. Similar to WT SCN neurons[7–10], the mean ± SEM input resistance of NT shRNA-expressing (black circles) SCN neurons was higher (P = 0.0001, one-way ANOVA) during the day than at night, and the mean ± SEM membrane potential was more depolarized (P = 0.004, one-way ANOVA), during the day than at night (Table 2). In contrast, there were no day–night differences (one-way ANOVA) in the mean ± SEM input resistances (D) or membrane potentials (E) of Kv12.1-targeted shRNA- (red circles) and Kv12.2-targeted shRNA-expressing (blue circles) SCN neurons (Table 2).
Acute in vivo shRNA-mediated knockdown of Kv12.1 or Kv12.2 increases the nighttime, but not the daytime, repetitive firing rates of SCN neurons. (A and B) Representative daytime (A) and nighttime (B) whole-cell current-clamp recordings obtained from NT shRNA- (black circles), Kv12.1-targeted shRNA- (red circles), and Kv12.2-targeted shRNA- (blue circles) expressing SCN neurons during the day (ZT7–ZT12) and at night (ZT18–ZT24) are shown. (C) Daytime and nighttime repetitive firing rates measured in individual SCN neurons expressing the NT (black circles), Kv12.1-targeted (red circles), or Kv12.2-targeted shRNA (blue circles) are plotted; mean ± SEM repetitive firing rates are indicated. Similar to WT SCN[7–10] neurons (Fig. 1), the mean ± SEM repetitive firing rate of NT shRNA-expressing (black circles) SCN neurons was lower (P = 0.0001, one-way ANOVA) at night than during the day (Table 2). In marked contrast, the mean ± SEM repetitive firing rates of Kv12.1- (red circles) and Kv12.2- (blue circles) targeted shRNA-expressing SCN neurons at night are not (one-way ANOVA) different from daytime firing rates (Table 2). (D and E) The input resistances (D) and membrane potentials (E) of NT shRNA- (black circles), Kv12.1-targeted shRNA- (red circles), and Kv12.2-targeted shRNA-expressing (blue circles) SCN neurons during the day and at night were also determined. Similar to WT SCN neurons[7–10], the mean ± SEM input resistance of NT shRNA-expressing (black circles) SCN neurons was higher (P = 0.0001, one-way ANOVA) during the day than at night, and the mean ± SEM membrane potential was more depolarized (P = 0.004, one-way ANOVA), during the day than at night (Table 2). In contrast, there were no day–night differences (one-way ANOVA) in the mean ± SEM input resistances (D) or membrane potentials (E) of Kv12.1-targeted shRNA- (red circles) and Kv12.2-targeted shRNA-expressing (blue circles) SCN neurons (Table 2).
Resting and active membrane properties of NT, Kv12.1-targeted, and Kv12.2-targeted shRNA-expressing SCN neurons during the day and at night
| Firing rate (Hz) | Rin (GΩ) | Vr (mV) | ISI (ms) | ||
|---|---|---|---|---|---|
| Day ZT 7–12 | NT shRNA | 3.9 ± 0.5* n = 24 | 2.1 ± 0.1 n = 23 | −41.5 ± 0.7** n = 24 | 221 ± 4* n = 24 |
| Kv12.1-targeted shRNA | 3.3 ± 0.5 n = 13 | 1.8 ± 0.2 n = 11 | −41.5 ± 1.2 n = 13 | 213 ± 9 n = 13 | |
| Kv12.2-targeted shRNA | 3.8 ± 0.6 n = 18 | 2.3 ± 0.3 n = 18 | −40.5 ± 0.9 n = 18 | 243 ± 8 n = 18 | |
| Night ZT 18–24 | NT shRNA | 1.6 ± 0.3* n = 27 | 0.9 ± 0.1* n = 22 | −45.8 ± 1.0** n = 27 | 366 ± 24* n = 27 |
| Kv12.1-targeted shRNA | 4.7 ± 0.7 n = 13 | 1.6 ± 0.2 n = 9 | −38.7 ± 1.9 n = 13 | 187 ± 5 n = 13 | |
| Kv12.2-targeted shRNA | 3.6 ± 0.8 n = 13 | 1.9 ± 0.3 n = 12 | −42.6 ± 1.5 n = 13 | 234 ± 6 n = 13 | |
Values are means ± SEM; n = number of cells; Rin = input resistance; Vr = resting membrane potentials; ISI = interspike interval. Values in daytime and nighttime NT shRNA-expressing neurons are significantly (*P = 0.0001; **P < 0.004) different.
Similar to the results obtained in WT, Kv12.1−/−, and Kv12.2−/− slices, the repetitive firing rates measured in daytime and nighttime NT, Kv12.1-targeted, and Kv12.2-targeted shRNA-expressing SCN neurons are quite variable. Similar variability in (in daytime and nighttime) repetitive firing rates was also evident across slices/animals, and comparisons of the averaged data acquired in individual slices (Fig. S4) revealed a marked (P = 0.01, one-way ANOVA) day–night difference in the mean repetitive firing rates measured in slices prepared from NT shRNA-expressing animals (Fig. S4 D and Table S1), whereas there were no day–night differences in mean repetitive firing rates determined in SCN slices prepared from Kv12.1-targeted (P = 0.18, one-way ANOVA) or Kv12.2-targeted (P = 0.64, one-way ANOVA) shRNA-expressing animals (Fig. S4 and Table S1).
Similar to WT SCN neurons (Table 1), input resistances were higher (P = 0.0001, one-way ANOVA), resting membrane potentials were more depolarized (P = 0.004, one-way ANOVA), and interspike intervals were shorter (P = 0.0001, one-way ANOVA) in NT shRNA-expressing SCN neurons during the day than at night (Fig. 3, D and E; and Table 2). In contrast, the input resistances, resting membrane potentials, and interspike intervals measured in Kv12.1-targeted and Kv12.2-targeted shRNA-expressing SCN neurons (P = 0.93, one-way ANOVA) at night and during the day were similar (Fig. 3, D and E; and Table 2). Similar results were obtained when the average data acquired in SCN slices from individual NT, Kv12.1-targeted, and Kv12.2-targeted shRNA-expressing SCN animals were compared (Fig. S4 and Table S1). The functional effects of the acute in vivo knockdown of Kv12.1 or Kv12.2 (Table 2 and Table S1) on the repetitive firing rates and membrane properties of SCN neurons, therefore, are virtually identical to those observed in Kv12.1−/− and Kv12.2−/− (Table 1 and Table S1) SCN neurons (see Discussion).
Kv12-encoded current densities are higher in nighttime, than in daytime, SCN neurons
The selective effects of the targeted deletion or the acute knockdown of Kv12.1 or Kv12.2 on the nighttime repetitive firing rates and membrane properties of SCN neurons suggest day–night differences in the functional expression of Kv12-encoded K+ currents. To explore this hypothesis directly, whole-cell voltage-clamp recordings were obtained from daytime and nighttime WT and DKO SCN neurons before and after the application of 20 μM CX4, reported to be a selective blocker of heterologously expressed Kv12.2-encoded K+ currents (Zhang et al., 2010). In these experiments, whole-cell outward K+ currents, evoked in response to action potentials waveforms (Fig. 4 A) obtained from nighttime WT SCN neurons, were recorded from daytime and nighttime WT (Fig. 4 B) and DKO (Fig. 4 C) SCN neurons with TTX and CdCl2 in the bath to block voltage-gated Na+ and Ca2+ channels, respectively. Offline digital subtraction of the currents recorded before and after exposure to CX4 provided the CX4-sensitive outward K+ currents. As is evident in the representative recordings shown in Fig. 4 B, the density of the CX4-sensitive K+ current is higher in the nighttime, compared with the daytime, WT SCN neuron. Similar results were obtained in recordings from multiple WT neurons (Fig. 4 D). The mean ± SEM CX4-sensitive current densities measured in daytime and nighttime WT SCN neurons were 7.9 ± 1.8 pA/pF (n = 17) and 23.6 ± 6.3 pA/pF (n = 13), respectively. The marked day–night difference in the densities of the CX4-sensitive outward K+ currents observed in WT SCN neurons, however, is not evident in DKO SCN neurons (Fig. 4 C). However, there are residual CX4-sensitive currents in DKO SCN neurons (Fig. 4 D), and the mean ± SEM CX4-sensitive outward K+ current densities measured in daytime and nighttime DKO SCN neurons were statistically indistinguishable at 4.3 ± 1.2 pA/pF (n = 8) and 5.6 ± 1.3 pA/pF (n = 8), respectively (Fig. 4 D). The densities of the residual CX4-sensitive outward K+ currents in daytime and nighttime DKO SCN neurons, therefore, are quite similar (see Discussion).

CX4-sensitive K + current densities in SCN neurons are higher at night than during the day. (A) Whole-cell outward K+ currents, evoked by an action potential voltage command recorded from a nighttime WT SCN neuron, were recorded in WT and DKO (Kv12.1−/−/Kv12.2−/−) SCN neurons before and after local application 20 μM CX4. Subtraction of the currents recorded before/after application of CX4 in each cell provided the CX4-sensitive K+ currents. (B and C) Representative CX4-sensitive outward K+ currents, obtained by offline digital subtraction of the currents recorded in the presence of CX4 from the currents recorded (in the same cell) in the control bath solution during the day and at night in WT (B) and DKO (C) SCN neurons, are shown. The CX4-sensitive outward K+ currents recorded in WT daytime and nighttime SCN neurons are also displayed on an expanded time scale in the insets in B. (D) Peak CX4-sensitive K+ current densities measured in WT (black squares) and DKO (magenta squares) SCN neurons during the day and at night are plotted. The mean ± SEM CX4-sensitive K+ current density in WT (black squares) SCN neurons is much higher (P = 0.002, one-way ANOVA) at night (n = 13) than during the day (n = 17). In contrast, CX4-sensitive K+ current densities in DKO (magenta squares) SCN neurons are low during the day (n = 8) and at night (n = 8), and there is no day–night difference (P = 0.85, one-way ANOVA) in mean ± SEM CX4-sensitive K+ current densities in DKO (magenta squares) SCN neurons.
CX4-sensitive K + current densities in SCN neurons are higher at night than during the day. (A) Whole-cell outward K+ currents, evoked by an action potential voltage command recorded from a nighttime WT SCN neuron, were recorded in WT and DKO (Kv12.1−/−/Kv12.2−/−) SCN neurons before and after local application 20 μM CX4. Subtraction of the currents recorded before/after application of CX4 in each cell provided the CX4-sensitive K+ currents. (B and C) Representative CX4-sensitive outward K+ currents, obtained by offline digital subtraction of the currents recorded in the presence of CX4 from the currents recorded (in the same cell) in the control bath solution during the day and at night in WT (B) and DKO (C) SCN neurons, are shown. The CX4-sensitive outward K+ currents recorded in WT daytime and nighttime SCN neurons are also displayed on an expanded time scale in the insets in B. (D) Peak CX4-sensitive K+ current densities measured in WT (black squares) and DKO (magenta squares) SCN neurons during the day and at night are plotted. The mean ± SEM CX4-sensitive K+ current density in WT (black squares) SCN neurons is much higher (P = 0.002, one-way ANOVA) at night (n = 13) than during the day (n = 17). In contrast, CX4-sensitive K+ current densities in DKO (magenta squares) SCN neurons are low during the day (n = 8) and at night (n = 8), and there is no day–night difference (P = 0.85, one-way ANOVA) in mean ± SEM CX4-sensitive K+ current densities in DKO (magenta squares) SCN neurons.
Although the action potential- (voltage-) clamp recordings (Fig. 4) reveal that the mean ± SEM CX4-sensitive outward K+ current density is considerably higher in nighttime than in daytime WT SCN neurons, there were a couple of daytime WT SCN neurons with higher than average CX4-sensitive current densities. In addition, the CX4-sensitive current densities measured in individual nighttime WT SCN neurons were quite variable (Fig. 4 D). To explore the functional consequences of these apparent differences, the effects of CX4 on the repetitive firing properties of SCN neurons were examined in whole-cell current-clamp recordings from cells in acute slices prepared from WT animals during the day (ZT7–ZT12) or at night (ZT18–ZT24). Consistent with the action potential- (voltage-) clamp data (Fig. 4), these experiments revealed that the application of 20 μM CX4 increased repetitive firing rates in only a small fraction (2 of 17; ∼10%) of daytime WT SCN neurons (Fig. 5 C), whereas the repetitive firing rates of most (11 of 15; ∼70%) nighttime WT SCN neurons were increased following application of 20 μM CX4 (Fig. 5 D). In addition to confirming day–night differences in the functional expression of Kv12-encoded K+ currents, these observations suggest that there is cellular heterogeneity in the subthreshold K+ channels mediating the day–night switch in the repetitive firing rates of SCN neurons (see Discussion).

Pairwise comparisons of repetitive firing rates in WT SCN neurons in the absence and presence of CX4. (A and B) Representative daytime (A) and nighttime (B) whole-cell current-clamp recordings obtained from WT SCN neurons before and after application of 20 μM CX4 are shown. (C and D) The repetitive firing rates measured in individual daytime (C) and nighttime (D) WT SCN neurons before and after application of 20 μM CX4 are plotted. In the vast majority (15 of 17; ∼90%) of WT SCN neurons, daytime repetitive firing rates were not affected (P = 0.6, repeated measures t test) by the application of 20 μM CX4 (C). In contrast, nighttime repetitive firing rates were increased (P = 0.001, repeated-measures Student’s t test) in most (11 of 15; ∼70%) WT SCN neurons upon application of 20 μM CX4 (D). Approximately 30% (4 of 15) of nighttime WT SCN neurons, however, were not measurably affected by 20 μM CX4.
Pairwise comparisons of repetitive firing rates in WT SCN neurons in the absence and presence of CX4. (A and B) Representative daytime (A) and nighttime (B) whole-cell current-clamp recordings obtained from WT SCN neurons before and after application of 20 μM CX4 are shown. (C and D) The repetitive firing rates measured in individual daytime (C) and nighttime (D) WT SCN neurons before and after application of 20 μM CX4 are plotted. In the vast majority (15 of 17; ∼90%) of WT SCN neurons, daytime repetitive firing rates were not affected (P = 0.6, repeated measures t test) by the application of 20 μM CX4 (C). In contrast, nighttime repetitive firing rates were increased (P = 0.001, repeated-measures Student’s t test) in most (11 of 15; ∼70%) WT SCN neurons upon application of 20 μM CX4 (D). Approximately 30% (4 of 15) of nighttime WT SCN neurons, however, were not measurably affected by 20 μM CX4.
Dynamic clamp-mediated subtraction of IKv12 increases firing rate in nighttime WT SCN neurons
To explore the impact of changing Kv12-encoded current amplitudes on the spontaneous repetitive firing rates of SCN neurons directly, we employed dynamic clamps to manipulate the Kv12-encoded current (IKv12) electronically in real time during current-clamp recordings. As described in Materials and methods, a model of IKv12 was developed using published data on heterologously expressed Kv12.1 currents (Engeland et al., 1998). The steady-state voltage-dependence of activation of IKv12 in the model was then adjusted to fit the voltage- (Fig. S3, A and B) and action potential- (Fig. 4) clamp data acquired from WT SCN neurons. The properties and amplitudes of IKv12 produced by the model (Fig. S3 C) reliably reproduce the CX4-sensitive currents identified in action potential-clamp recordings from WT SCN neurons.
In all dynamic clamp experiments, whole-cell current-clamp recordings were obtained from SCN neurons in acute slices prepared from adult WT animals at night or during the day. After recording gap-free spontaneous firing activity under control conditions (Fig. 6, A and E), the effects on spontaneous repetitive firing rates of subtracting or increasing IKv12 amplitudes were determined. Initial experiments were conducted on nighttime WT SCN neurons to determine the minimal subtracted IKv12 amplitude that increased the spontaneous repetitive firing rates of most (9 of 10) cells. These experiments revealed that the minimal (peak) amplitude of IKv12 amplitude that increased the repetitive firing rates of most (9 of 10) nighttime WT SCN neurons was 2 pA. This value (2 pA) was then set equal to x, and the effects of subtracting varying amplitudes (in integral multiples of x) of IKv12 on the repetitive firing rates of WT nighttime neurons were determined. For daytime WT neurons, IKv12 was scaled (to 0.5 pA) to reflect the day–night difference in the (mean) CX4-sensitive current amplitudes measured in WT neurons (Fig. 4 D), and the effects of the subtraction of integral multiples of this value (i.e., x = 0.5 pA) on the repetitive firing rates of daytime WT neurons were determined. As shown in the representative recordings from a WT nighttime SCN neuron in Fig. 6, A–C, subtraction of the minimal IKv12 (−x) increased the rate of repetitive firing (Fig. 6 B), and the rate was increased further when the amplitude of IKv12 was increased (−3x) threefold (Fig. 6 C). In contrast, as illustrated in the representative recordings from a WT daytime SCN neuron in Fig. 6, E–G, dynamic clamp-mediated subtraction of IKv12 (at −x or −3x) had little to no effect on the rate of spontaneous action potential firing. Similar results were obtained in recordings from additional WT nighttime and daytime SCN neurons in which varying amplitudes (−x, −2x, −3x, −4x, and −5x) of IKv12 were subtracted via a dynamic clamp. The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT daytime (open black square; n = 7) and nighttime (closed black square; n = 10) SCN neurons are plotted as a function of the magnitude of IKv12 subtracted in Fig. 6, D and H. The insets below Fig. 6, C and G, show the waveforms of individual action potentials recorded in the representative WT nighttime (C) and daytime (G) SCN neurons, plotted on an expanded timescale. The modeled (−3x) IKv12 waveforms generated in these cells are shown below the voltage records; the zero current levels are indicated by dotted lines. It is of interest to note that IKv12 increases as the membrane potential approaches the threshold for generating an action potential in the nighttime neuron (Fig. 6 C, inset), and the mean modeled IKv12 amplitudes are larger during interspike intervals in nighttime (1.49 ± 0.15 pA; Fig. 6 C, inset) than in daytime (−0.39 ± 0.05 pA; Fig. 6 G, inset) SCN neurons. These observations are consistent with a selective role for IKv12 in regulating the interspike interval and the rate of spontaneous repetitive action potential firing in nighttime SCN neurons.

Dynamic clamp-mediated subtraction of I Kv12 increases the repetitive firing rates of nighttime WT SCN neurons. (A–C, and E–G) Representative whole-cell current-clamp recordings from a WT nighttime (A–C) and a WT daytime (E–G) SCN neuron at baseline (A and E) and with different magnitudes of modeled IKv12 subtracted (B, C, F, and G) via dynamic clamp are shown. In the insets below C and G, the waveforms of individual action potentials (black) recorded in the representative WT nighttime (C) and daytime (G) SCN neurons are displayed and plotted on an expanded timescale. Below these voltage records, the modeled (−3x) IKv12 waveforms (purple) are shown; the zero current levels are indicated by the dotted lines. As is evident, the effect of subtracting increasing IKv12 is greater in the nighttime (B and C), than in the daytime (F and G), WT SCN neuron. Similar results were obtained in current-clamp recordings from additional WT nighttime and daytime SCN neurons in which varying amplitudes (−x, −2x, −3x, −4x, and −5x) of the minimal modeled IKv12 (0.5 pA) were subtracted via dynamic clamp. (D and H) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT daytime (empty square; n = 10) and nighttime (filled square; n = 7) SCN neurons are plotted as a function of the magnitude of modeled IKv12 subtracted.
Dynamic clamp-mediated subtraction of I Kv12 increases the repetitive firing rates of nighttime WT SCN neurons. (A–C, and E–G) Representative whole-cell current-clamp recordings from a WT nighttime (A–C) and a WT daytime (E–G) SCN neuron at baseline (A and E) and with different magnitudes of modeled IKv12 subtracted (B, C, F, and G) via dynamic clamp are shown. In the insets below C and G, the waveforms of individual action potentials (black) recorded in the representative WT nighttime (C) and daytime (G) SCN neurons are displayed and plotted on an expanded timescale. Below these voltage records, the modeled (−3x) IKv12 waveforms (purple) are shown; the zero current levels are indicated by the dotted lines. As is evident, the effect of subtracting increasing IKv12 is greater in the nighttime (B and C), than in the daytime (F and G), WT SCN neuron. Similar results were obtained in current-clamp recordings from additional WT nighttime and daytime SCN neurons in which varying amplitudes (−x, −2x, −3x, −4x, and −5x) of the minimal modeled IKv12 (0.5 pA) were subtracted via dynamic clamp. (D and H) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT daytime (empty square; n = 10) and nighttime (filled square; n = 7) SCN neurons are plotted as a function of the magnitude of modeled IKv12 subtracted.
In contrast with the marked differences in the effects of subtracting IKv12 in WT nighttime and daytime SCN neurons (Fig. 6), dynamic clamp-mediated addition of IKv12 decreased the repetitive firing rates of both nighttime and daytime WT SCN neurons (Fig. 7). The addition of the modeled IKv12 also reduced the rate of repetitive firing in nighttime Kv12.1−/− SCN neurons (Fig. S6, A–C), although the magnitudes of the percent changes in the repetitive firing rates of nighttime Kv12.1−/− SCN neurons were smaller than in nighttime WT SCN neurons (Fig. S6 D).

Dynamic clamp-mediated addition of modeled Kv12-encoded (I Kv12 ) currents in WT SCN neurons. (A–C, and E–G) Representative whole-cell current-clamp recordings from a WT nighttime (A–C) and a WT daytime (E−G) SCN neuron at baseline (A and E) and with different magnitudes of modeled IKv12 added (B, C, F, and G) via dynamic clamp are shown. In the insets below C and G, the waveforms of individual action potentials (black) recorded in the representative WT nighttime (C) and daytime (G) SCN neurons are displayed and plotted on an expanded timescale. Below these voltage records, the modeled (+16x) IKv12 waveforms (purple) are shown; the dotted lines indicate the zero current levels. As is evident, increasing IKv12 reduced the rate of repetitive firing in both the WT nighttime (B and C) and the WT daytime (F and G) SCN neuron, although the impact is greater on nighttime (B and C) than on daytime (F and G) firing. Similar results were obtained in recordings from additional WT nighttime and daytime SCN neurons in which varying amplitudes (+x, +2x, +4x, +8x, and +16x) of the minimal modeled IKv12 (2 pA) were added during current-clamp recordings. (D and H) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT daytime (empty circles, n = 7) and nighttime (filled circles, n = 10) SCN neurons are plotted as a function of the magnitude of modeled IKv12 added.
Dynamic clamp-mediated addition of modeled Kv12-encoded (I Kv12 ) currents in WT SCN neurons. (A–C, and E–G) Representative whole-cell current-clamp recordings from a WT nighttime (A–C) and a WT daytime (E−G) SCN neuron at baseline (A and E) and with different magnitudes of modeled IKv12 added (B, C, F, and G) via dynamic clamp are shown. In the insets below C and G, the waveforms of individual action potentials (black) recorded in the representative WT nighttime (C) and daytime (G) SCN neurons are displayed and plotted on an expanded timescale. Below these voltage records, the modeled (+16x) IKv12 waveforms (purple) are shown; the dotted lines indicate the zero current levels. As is evident, increasing IKv12 reduced the rate of repetitive firing in both the WT nighttime (B and C) and the WT daytime (F and G) SCN neuron, although the impact is greater on nighttime (B and C) than on daytime (F and G) firing. Similar results were obtained in recordings from additional WT nighttime and daytime SCN neurons in which varying amplitudes (+x, +2x, +4x, +8x, and +16x) of the minimal modeled IKv12 (2 pA) were added during current-clamp recordings. (D and H) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT daytime (empty circles, n = 7) and nighttime (filled circles, n = 10) SCN neurons are plotted as a function of the magnitude of modeled IKv12 added.

Dynamic clamp-mediated addition of I Kv12 decreases the rate of repetitive firing in nighttime Kv12.1 −/− SCN neurons. (A–C) Representative whole-cell current-clamp recordings obtained from a nighttime (ZT18–ZT20) Kv12.1−/− SCN neuron under control conditions (A) and with dynamic clamp-mediated addition of modeled IKv12 (B and C). Similar to WT SCN neurons (see Fig. 7), the addition of modeled IKv12 (B and C) reduced the repetitive firing rates of Kv12.1−/− SCN neurons in direct proportion to the amplitude of the injected current. (D) The mean ± SEM (n = 11) percentage changes in the repetitive firing rates of nighttime Kv12.1−/− SCN neurons (red circles) are plotted as a function of the amplitude of IKv12 added. The percent changes in the repetitive firing rates of WT neurons with the addition of IKv12 (from Fig. 7 D) are replotted here (black circles) to facilitate direct comparison of the results in WT and Kv12.1−/− SCN neurons.
Dynamic clamp-mediated addition of I Kv12 decreases the rate of repetitive firing in nighttime Kv12.1 −/− SCN neurons. (A–C) Representative whole-cell current-clamp recordings obtained from a nighttime (ZT18–ZT20) Kv12.1−/− SCN neuron under control conditions (A) and with dynamic clamp-mediated addition of modeled IKv12 (B and C). Similar to WT SCN neurons (see Fig. 7), the addition of modeled IKv12 (B and C) reduced the repetitive firing rates of Kv12.1−/− SCN neurons in direct proportion to the amplitude of the injected current. (D) The mean ± SEM (n = 11) percentage changes in the repetitive firing rates of nighttime Kv12.1−/− SCN neurons (red circles) are plotted as a function of the amplitude of IKv12 added. The percent changes in the repetitive firing rates of WT neurons with the addition of IKv12 (from Fig. 7 D) are replotted here (black circles) to facilitate direct comparison of the results in WT and Kv12.1−/− SCN neurons.
To determine if the functional effects of dynamic clamp-mediated manipulation of IKv12 observed in WT SCN neurons (Figs. 6 and 7) are specific for IKv12 or, alternatively, are observed when any subthreshold voltage-activated K+ current is added/subtracted, we generated a mathematical model of IA in SCN neurons. In previous studies, we have shown that shRNA-mediated knockdown of Kv4.1 decreases IA amplitudes and increases the repetitive firing rates of daytime and nighttime SCN neurons (Hermanstyne et al., 2017). Using acquired voltage-clamp data describing the time- and voltage-dependent properties of IA in WT SCN neurons (Hermanstyne et al., 2017), we generated a model of IA gating (Fig. S3). As illustrated in Fig. S7, the modeled IA waveforms and IKv12 waveforms generated by individual action potentials recorded in a representative WT SCN neuron are distinct.

Kinetic differences between the Cybercyte modeled I A and I Kv12 . Comparison of the normalized currents produced by the Cybercyte IKv12 (lower panel, purple) and IA (lower panel, blue) models generated by action potential waveforms (upper panel, black) from a representative WT SCN neuron.
Kinetic differences between the Cybercyte modeled I A and I Kv12 . Comparison of the normalized currents produced by the Cybercyte IKv12 (lower panel, purple) and IA (lower panel, blue) models generated by action potential waveforms (upper panel, black) from a representative WT SCN neuron.
In experiments similar to those described above for IKv12, we determined the effects on spontaneous repetitive rates of subtracting/adding IA, presented in multiples of the minimal IA, x, determined as described in Materials and methods. Dynamic clamp-mediated subtraction of modeled IA initially increased the repetitive firing rates of WT nighttime (Fig. 8, A, C, and E) and daytime (Fig. 8, B, D, and F) SCN neurons. With the subtraction of higher amplitude currents, however, repetitive firing rates decreased in WT nighttime (Fig. 8 E) and daytime (Fig. 8 F) SCN neurons. The insets below Fig. 8, C and D, show the waveforms of individual action potentials recorded in the representative WT nighttime (C) and daytime (D) SCN neurons, plotted on an expanded timescale. The modeled (−3x) IA waveforms generated in these cells are shown below the voltage records: the 0 current levels are indicated by dotted lines. As is evident, there are clear differences in the peak amplitudes of the modeled IA during the action potentials in WT nighttime (C) and daytime (D) SCN neurons. In addition and in contrast with the results obtained with IKv12 (Fig. 6), IA is prominent during interspike intervals in WT nighttime (−6.88 ± 0.83 pA; Fig. 8 C, inset) and daytime (−7.03 ± 0.34 pA; Fig. 8 D, inset) SCN neurons, an observation consistent with the physiological role of IA in driving the repetitive firing rates of SCN neurons during the day and at night (Hermanstyne et al., 2017). The addition of modeled IA also decreased the spontaneous repetitive firing rates of WT nighttime (Fig. 9, A, C, and E) and daytime (Fig. 9, B, D, and F) SCN neurons similarly.

Dynamic clamp-mediated subtraction of modeled I A in WT SCN neurons. (A–D) Representative whole-cell current-clamp recordings from WT nighttime (A and C) and daytime (B and D) SCN neurons with different magnitudes of modeled IA subtracted are shown. In the insets below C and D, the waveforms of individual action potentials (black), recorded in the representative WT nighttime (C) and daytime (D) SCN neurons, are plotted on an expanded timescale. The modeled (−3x) IKv12 waveforms (blue) generated in these cells are shown; the zero current levels are indicated by the dotted lines. As is evident, the rates of repetitive firing of the representative nighttime (A and C) and daytime (B and D) SCN neurons are increased with the subtraction of modeled IA. Similar results were obtained in recordings from additional nighttime and daytime WT SCN in which varying amplitudes of the minimal modeled IA (20 pA) were subtracted (−x, −2x, −3x, −4x, and −5x) during current-clamp recordings. (E and F) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT SCN neurons in response to subtracting (open and closed squares; n = 16) modeled IA are plotted as a function of the magnitude of the modeled IA subtracted.
Dynamic clamp-mediated subtraction of modeled I A in WT SCN neurons. (A–D) Representative whole-cell current-clamp recordings from WT nighttime (A and C) and daytime (B and D) SCN neurons with different magnitudes of modeled IA subtracted are shown. In the insets below C and D, the waveforms of individual action potentials (black), recorded in the representative WT nighttime (C) and daytime (D) SCN neurons, are plotted on an expanded timescale. The modeled (−3x) IKv12 waveforms (blue) generated in these cells are shown; the zero current levels are indicated by the dotted lines. As is evident, the rates of repetitive firing of the representative nighttime (A and C) and daytime (B and D) SCN neurons are increased with the subtraction of modeled IA. Similar results were obtained in recordings from additional nighttime and daytime WT SCN in which varying amplitudes of the minimal modeled IA (20 pA) were subtracted (−x, −2x, −3x, −4x, and −5x) during current-clamp recordings. (E and F) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT SCN neurons in response to subtracting (open and closed squares; n = 16) modeled IA are plotted as a function of the magnitude of the modeled IA subtracted.

Dynamic clamp-mediated addition of modeled I A in WT SCN neurons. (A–D) Representative whole-cell current-clamp recordings from WT nighttime (A and C) and daytime (B and D) SCN neurons with different magnitudes of modeled IA added are shown. In the insets below C and D, the waveforms of individual action potentials (black), recorded in the representative WT nighttime (C) and daytime (D) SCN neurons, are plotted on an expanded timescale. The modeled (+3x) IKv12 waveforms (blue) generated in these cells are shown; the zero current levels are indicated by the dotted lines. The rates of repetitive firing of both the representative nighttime (A and C) and the representative daytime (B and D) SCN neurons are decreased with the addition of modeled IA. Recordings from additional nighttime and daytime WT SCN neurons in which varying amplitudes (x, 2x, 3x, 4x, 5x) of the minimal modeled IA (20 pA) were added also revealed a decrease in repetitive firing rates. (E–H) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT SCN neurons in response to adding (open and closed circles; n = 14) modeled IA are plotted as a function of the magnitude of the modeled IA added.
Dynamic clamp-mediated addition of modeled I A in WT SCN neurons. (A–D) Representative whole-cell current-clamp recordings from WT nighttime (A and C) and daytime (B and D) SCN neurons with different magnitudes of modeled IA added are shown. In the insets below C and D, the waveforms of individual action potentials (black), recorded in the representative WT nighttime (C) and daytime (D) SCN neurons, are plotted on an expanded timescale. The modeled (+3x) IKv12 waveforms (blue) generated in these cells are shown; the zero current levels are indicated by the dotted lines. The rates of repetitive firing of both the representative nighttime (A and C) and the representative daytime (B and D) SCN neurons are decreased with the addition of modeled IA. Recordings from additional nighttime and daytime WT SCN neurons in which varying amplitudes (x, 2x, 3x, 4x, 5x) of the minimal modeled IA (20 pA) were added also revealed a decrease in repetitive firing rates. (E–H) The mean ± SEM percent changes in the spontaneous repetitive firing rates of WT SCN neurons in response to adding (open and closed circles; n = 14) modeled IA are plotted as a function of the magnitude of the modeled IA added.
Additional experiments were completed to examine the effects of subtracting modeled IKv12 versus modeled IA in the same daytime cell. As in the experiments presented in Fig. 8, the subtraction of IA initially increased repetitive firing rates (Fig. S8, A and B), and at higher current amplitudes, firing became unreliable (Fig. S8 C). In contrast, subtraction of IKv12 in the same cell had very little effect on repetitive firing rates (Fig. S8, D–F), with results similar to those presented in Fig. 6. The functional effects of subtracting modeled IA versus modeled IKv12 on the repetitive firing rates of SCN neurons, therefore, are distinct.

Direct comparison of the effects of dynamic clamp-mediated subtraction of I Kv12 versus I A in daytime WT SCN neurons. (A, B, D, and E) Representative whole-cell current-clamp recordings were obtained from a day-phased (ZT7–ZT12) WT SCN neuron with dynamic clamp-mediated subtraction of modeled IA (A and B) or IKv12 (D and E). Subtracting modeled 1x or 3x IA increased the rate of repetitive firing (A and B), whereas subtracting modeled 1x or 3x IKv12 (D and E) in the same cell did not measurably alter the firing rate. In the insets below B and E, the waveforms of individual action potentials (black), recorded in a representative WT daytime SCN neuron with subtracted modeled (−3x) IA (B) or (−3x) IKv12 (E), are plotted on an expanded timescale. The modeled (−3x) IA (blue, B) and (−3x) IKv12 (purple, E) waveforms in these cells are shown; the zero current levels are indicated by the dotted lines. Similar results were obtained in six additional cells. (C and F) Plotting the mean ± SEM (n = 7) percentage changes in the repetitive firing rates of daytime WT SCN neurons reveals the mark difference in the effects of subtracting IA (C) versus IKv12 (F).
Direct comparison of the effects of dynamic clamp-mediated subtraction of I Kv12 versus I A in daytime WT SCN neurons. (A, B, D, and E) Representative whole-cell current-clamp recordings were obtained from a day-phased (ZT7–ZT12) WT SCN neuron with dynamic clamp-mediated subtraction of modeled IA (A and B) or IKv12 (D and E). Subtracting modeled 1x or 3x IA increased the rate of repetitive firing (A and B), whereas subtracting modeled 1x or 3x IKv12 (D and E) in the same cell did not measurably alter the firing rate. In the insets below B and E, the waveforms of individual action potentials (black), recorded in a representative WT daytime SCN neuron with subtracted modeled (−3x) IA (B) or (−3x) IKv12 (E), are plotted on an expanded timescale. The modeled (−3x) IA (blue, B) and (−3x) IKv12 (purple, E) waveforms in these cells are shown; the zero current levels are indicated by the dotted lines. Similar results were obtained in six additional cells. (C and F) Plotting the mean ± SEM (n = 7) percentage changes in the repetitive firing rates of daytime WT SCN neurons reveals the mark difference in the effects of subtracting IA (C) versus IKv12 (F).
Kcnh8 and Kcnh3 transcript expression in the SCN do not display 24-h rhythms
The results of the action potential- (voltage-) clamp experiments (Fig. 4) demonstrate that amplitudes/densities of the CX4-sensitive, Kv12-encoded K+ currents in SCN neurons are higher at night than during the day, suggesting that the functional expression of the underlying K+ channels is regulated by the molecular clock. To determine if this regulation reflects day–night differences in the expression levels of the Kcnh transcripts encoding the Kv12.1 (Kcnh8) and Kv12.2 (Kcnh3) α subunits, quantitative real-time PCR (qRT-PCR) analysis was performed on RNA samples extracted from WT SCN (n = 7–8) every 4 h over two consecutive days. The expression levels of the transcripts encoding the clock genes, Per2 and Bmal (Reppert and Weaver, 2001; Hastings et al., 2018, 2019; Michel and Meijer, 2020), and of Hprt (control gene) in every sample were also determined. Expression of the Per2, Bmal, Kcnh8, and Kcnh3 transcripts was normalized to Hprt in the same sample and evaluated for rhythmicity using JTK cycle analysis (Hughes et al., 2010) with the period set to 24 h. As expected, these analyses revealed that Per2 and Bmal display ∼24 h anti-phase rhythms (Fig. S9) in expression (Reppert and Weaver, 2001; Hastings et al., 2018, 2019; Michel and Meijer, 2020). In contrast, the mRNA expression levels of Kcnh8 and Kcnh3 do not (P > 0.05, JTK cycle Bonferroni-adjusted) display 24-h rhythms (Fig. S8), indicating that oscillations in the expression of the Kcnh8 and/or Kcnh3 transcripts do not underlie the observed day–night differences in the functional expression of Kv12-encoded currents (see Discussion).

Expression of Kcnh8 and Kcnh3 in the SCN. The expression levels of the transcripts encoding Per2, Bmal, Kcnh8, and Kcnh3 were determined by quantitative RT-PCR analyses of SCN tissues samples, obtained every 4 h over a 48 h time-period, from animals maintained in the standard and reversed 12:12 h LD conditions, as described in Materials and methods. The expression of each transcript was normalized to the expression of Hprt in the same sample. Mean ± SEM (n = 7–8) values for each transcript are plotted. Fitting the mean data for each transcript, using JTK cycle analysis (Hughes et al., 2010) with the period set to 24 h, reveals that, as expected (Kuhlman and McMahon, 2006; Hastings et al., 2019), the Per2 (P = 0.0001) and Bmal (P = 0.0001) transcripts display 24 rhythms in expression. In contrast, the expression levels of the Kcnh8 (P = 1.00) and Kcnh3 (P = 0.102) transcripts do not display 24 h rhythms. The dashed lines on days 1 and 2 are at 7 pm.
Expression of Kcnh8 and Kcnh3 in the SCN. The expression levels of the transcripts encoding Per2, Bmal, Kcnh8, and Kcnh3 were determined by quantitative RT-PCR analyses of SCN tissues samples, obtained every 4 h over a 48 h time-period, from animals maintained in the standard and reversed 12:12 h LD conditions, as described in Materials and methods. The expression of each transcript was normalized to the expression of Hprt in the same sample. Mean ± SEM (n = 7–8) values for each transcript are plotted. Fitting the mean data for each transcript, using JTK cycle analysis (Hughes et al., 2010) with the period set to 24 h, reveals that, as expected (Kuhlman and McMahon, 2006; Hastings et al., 2019), the Per2 (P = 0.0001) and Bmal (P = 0.0001) transcripts display 24 rhythms in expression. In contrast, the expression levels of the Kcnh8 (P = 1.00) and Kcnh3 (P = 0.102) transcripts do not display 24 h rhythms. The dashed lines on days 1 and 2 are at 7 pm.
Loss of Kv12 channels does not eliminate rhythms in (wheel-running) locomotor activity
Representative recordings of wheel-running activity in WT, Kv12.1−/−, and Kv12.2−/− mice are presented in Fig. 10 A. In these experiments, all mice were entrained to a standard 12:12 h LD cycle for 10 d, followed by at least 20 d in DD (Fig. 10 A). Circadian periods were measured in each animal after 20 d in DD (Fig. 10 C). The mean ± SEM circadian periods determined in adult WT (23.6 ± 0.1 h; n = 9), Kv12.1−/− (23.5 ± 0.1 h; n = 15), Kv12.2−/− (23.6 ± 0.1 h; n = 14), and DKO (23.4 ± 0.1 h; n = 12, data not shown) animals are indistinguishable (Fig. 10 C). Additional experiments conducted on WT mice 2 wk following bilateral injections of the Kv12.1-targeted, Kv12.2-targeted, or NT-shRNA-expressing AAV8 into the SCN revealed that the acute in vivo knockdown of Kv12.1 or Kv12.2 in the adult SCN also did not disrupt rhythmic wheel-running activity (Fig. 10, B and D).

Rhythmic wheel-running activity is maintained in Kv12.1 −/− and Kv12.2 −/− mice and with acute in vivo shRNA-mediated knockdown of Kv12.1 or Kv12.2 in adult animals. (A and B) Representative recordings of wheel-running activity of WT (top), Kv12.1−/− (middle), and Kv12.2−/− (bottom) mice (A) and WT mice that received bilateral SCN injections of the non-targeted (top), Kv12.1-targeted (middle), or Kv12.2-targeted (bottom) shRNA-expressing AAV8 (B). Continuous recordings were obtained for ∼10 d in 12:12 h LD (indicated by the grey and white backgrounds, respectively) conditions, followed by at least 20 d in DD (constant darkness, indicated by the white background). The circadian periods, measured in each WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) mouse and in each animal expressing the NT (black circles), Kv12.1-targeted (red circles) and Kv12.2-targeted (blue circles) shRNA in DD, are plotted; P values (one-way ANOVA are indicated). (C) The mean ± SEM circadian periods of locomotor activity determined in WT (black squares; n = 9), Kv12.1−/− (red squares; n = 15) and Kv12.2−/− (blue squares; n = 14) are similar. (D) The mean ± SEM circadian periods (D) of locomotor activity measured in animals expressing the Kv12.1-targeted shRNA (red circles; n = 7), the Kv12.2-targeted shRNA (blue circles; n = 7) and the NT shRNA (black circles; n = 11) are also similar.
Rhythmic wheel-running activity is maintained in Kv12.1 −/− and Kv12.2 −/− mice and with acute in vivo shRNA-mediated knockdown of Kv12.1 or Kv12.2 in adult animals. (A and B) Representative recordings of wheel-running activity of WT (top), Kv12.1−/− (middle), and Kv12.2−/− (bottom) mice (A) and WT mice that received bilateral SCN injections of the non-targeted (top), Kv12.1-targeted (middle), or Kv12.2-targeted (bottom) shRNA-expressing AAV8 (B). Continuous recordings were obtained for ∼10 d in 12:12 h LD (indicated by the grey and white backgrounds, respectively) conditions, followed by at least 20 d in DD (constant darkness, indicated by the white background). The circadian periods, measured in each WT (black squares), Kv12.1−/− (red squares), and Kv12.2−/− (blue squares) mouse and in each animal expressing the NT (black circles), Kv12.1-targeted (red circles) and Kv12.2-targeted (blue circles) shRNA in DD, are plotted; P values (one-way ANOVA are indicated). (C) The mean ± SEM circadian periods of locomotor activity determined in WT (black squares; n = 9), Kv12.1−/− (red squares; n = 15) and Kv12.2−/− (blue squares; n = 14) are similar. (D) The mean ± SEM circadian periods (D) of locomotor activity measured in animals expressing the Kv12.1-targeted shRNA (red circles; n = 7), the Kv12.2-targeted shRNA (blue circles; n = 7) and the NT shRNA (black circles; n = 11) are also similar.
Discussion
The results of the experiments detailed here demonstrate that the targeted deletion of Kv12.1 or Kv12.2 or the shRNA-mediated knockdown of Kv12.1 or Kv12.2 selectively increased the repetitive firing rates of SCN neurons at night and eliminated the day–night difference in mean repetitive firing rates that is characteristic of WT SCN neurons (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020). Direct application of the Kv12 channel blocker CX4 (Zhang et al., 2010) increased repetitive firing rates at night, but not during the day, and the amplitudes of CX4-sensitive currents in WT SCN neurons were found to be higher at night than during the day. Additional experiments revealed that dynamic clamp-mediated subtraction of modeled IKv12 selectively increased the repetitive firing rates of nighttime, and not daytime, SCN neurons. Despite the loss of day–night oscillations in mean repetitive firing rates, mice lacking Kv12.1 or/and Kv12.2 currents display daily rhythms in locomotor (wheel-running) activity.
Kv12 channels selectively regulate the nighttime repetitive firing properties of SCN neurons
The targeted deletion of either Kcnh3 (Kv12.2) or Kcnh8 (Kv12.1) selectively increased the rate of spontaneous action potential firing of SCN neurons at night; the daytime repetitive firing rates of SCN neurons lacking Kv12.1 or Kv12.2 are indistinguishable from WT cells. The higher repetitive firing rates in nighttime Kv12.1−/− and Kv12.2−/− compared with WT, SCN neurons are accompanied by higher input resistances and depolarized membrane potentials. The selective effects on nighttime firing rates and membrane properties were also observed in DKO SCN neurons lacking both Kv12.1 and Kv12.2 (Table 1). Acute, in vivo Kv12.1-targeted or Kv12.2-targeted shRNA-mediated knockdown of Kv12.1 or Kv12.2 in adult SCN neurons also selectively increased nighttime repetitive firing rates (Table 2). In addition to demonstrating a critical physiological role for Kv12-encoded K+ channels in regulating the nighttime firing and membrane properties of mature SCN neurons, these results also suggest that the firing properties of Kv12.1−/−, Kv12.2−/−, and Kv12.1−/−/Kv12.2−/− SCN neurons do not reflect effects secondary to the loss of Kv12.1/Kv12.2 channels throughout development. Mimicking the effects of knockout/knockdown of Kv12.1/Kv12.2, dynamic clamp-mediated subtraction of IKv12 selectively increased the repetitive firing rates of WT nighttime SCN neurons. These results contrast with the effects of manipulating another subthreshold K+ current, IA: dynamic clamp-mediated subtraction of IA increased spontaneous repetitive firing rates in WT nighttime and daytime SCN neurons.
Application of 20-μM CX4 also selectively increased the spontaneous repetitive firing rates of nighttime WT SCN neurons, and voltage-clamp experiments revealed that CX4-sensitive outward K+ currents are higher in nighttime than in daytime WT SCN neurons. In the original study characterizing CX4 (Zhang et al., 2010), however, it was reported that CX4 is a selective blocker of heterologously expressed Kv12.2-encoded K+ currents and that expressed Kv12.1-encoded K+ currents were unaffected by CX4. The finding here that application of 20 μM CX4 markedly increases the repetitive firing rates of nighttime SCN neurons, similar to the knockout/knockdown of either Kv12.1 or Kv12.2, suggests that CX4 may also affect Kv12.1-encoded K+ channels in native (neuronal) cells. Alternatively, these results may suggest that Kv12.1 and Kv12.2 form heteromeric channels in SCN neurons. The presence of the residual CX-sensitive outward K+ currents in DKO SCN neurons (Fig. 4) reveals another caveat with the interpretation of these pharmacological experiments, i.e., there is a component(s) of the CX4-sensitive outward K+ currents that reflects the effects of CX4 on non-Kv12-encoded K+ channels. Additional experiments will be needed to identify the molecular determinant(s) of the residual, non-Kv12-encoded component of the CX4-insensitive currents in SCN neurons.
Relationship to previous studies
Several previous studies have demonstrated selective roles for other (i.e., non-Kv12-encoded) types of K+ channels in regulating the daytime and/or nighttime repetitive firing rates of SCN neurons. Acute in vivo shRNA-mediated knockdown of Kcnd1, which encodes the Kv4.1 α subunit and contributes to the generation of IA, in adult SCN neurons affects daytime and nighttime IA densities and spontaneous firing rates (Hermanstyne et al., 2017). The densities of IA were lower in Kv4.1-targeted shRNA-expressing compared with WT, SCN neurons during the day and at night (Hermanstyne et al., 2017). Knockdown of Kv4.1 also increased repetitive firing rates during the day and at night (Hermanstyne et al., 2017). In contrast with the effects of the knockdown or deletion of Kv12 channels, the loss of Kv4.1-encoded IA channels did not eliminate day–night oscillations in the repetitive firing rates of SCN neurons (Hermanstyne et al., 2017). Also in contrast with the results here, in vivo knockdown of Kv4.1-encoded IA channels advanced the time of daily activity onset by ∼1.8 h and shortened the periods of wheel-running activity in constant darkness by ∼0.5 h (Hermanstyne et al., 2017).
In contrast with the effects of manipulating IA (Granados-Fuentes et al., 2012, 2015; Hermanstyne et al., 2017), targeted deletion of Kcnma1, which encodes the pore-forming α subunit of large conductance voltage- and Ca2+-dependent K+ (BK) channels, results in increased repetitive firing rates selectively in nighttime SCN neurons (Meredith et al., 2006), markedly reducing the day–night oscillation in mean repetitive firing rates, characteristic of WT SCN neurons (Brown and Piggins, 2007; Allen et al., 2017; Belle and Allen, 2018; Harvey et al., 2020). The circadian periods of locomotor activity and clock gene expression in Kcnma1−/− animals, however, were indistinguishable from WT animals, although the circadian amplitudes of wheel-running activity and other behavioral outputs were reduced in Kcnma1−/−, compared with WT, mice during the day and at night (Meredith et al., 2006). Opposite cellular effects were seen with overexpression of Kcnma1 in the SCN, i.e., the repetitive firing rates were reduced in the daytime but not in the nighttime SCN neurons (Montgomery et al., 2013). The overexpression of Kcnma1 and the reduction in daytime repetitive firing rates, however, were not accompanied by changes in the periods or the amplitudes of circadian behaviors (Montgomery et al., 2013).
Previous studies have also demonstrated marked day–night differences in fast delayed rectifier (FDR) K+ current densities and in the expression of the Kcnc1 and Kcnc2 transcripts, which encode the Kv3.1b and Kv3.2 α subunits and generate FDR K+ currents, in SCN neurons (Itri et al., 2005). In addition, it was reported that exposure to the K+ channel blocker, 4-aminopyridine (4-AP) at a concentration of 0.5 mM, selectively reduces daytime repetitive firing rates and eliminates daily rhythms in the firing rates of SCN neurons (Itri et al., 2005). Similar effects on daytime firing rates were seen in SCN neurons from (Kcnc1−/−/Kcnc2−/−) animals lacking both Kv3.1b and Kv3.2 (Kudo et al., 2011), consistent with the suggestion that 0.5 mM 4-AP selectively affects FDR K+ currents in SCN neurons (Itri et al., 2005). Despite the elimination of day–night oscillations in repetitive firing rates, daily rhythms in Per2 expression were intact in Kcnc1−/−/Kcnc2−/− animals (Kudo et al., 2011).
Molecular determinants of day–night differences in Kv12-encoded K+ current densities in the SCN
Consistent with the selective effects of the deletion or the acute knockdown of Kv12 subunits, action potential- (voltage-) clamp experiments revealed that the mean density of the CX4-sensitive (Kv12-encoded) K+ currents in WT SCN neurons is much higher at night than during the day. The quantitative RT-PCR analysis completed, however, indicates that the day–night difference in Kv12-encoded K+ current densities does not result from circadian regulation of the Kcnh3/Kcnh8 transcripts encoding the Kv12.1/Kv12.2 α subunits. Although it is certainly possible that Kv12.1/Kv12.2 protein expression in the SCN is rhythmic despite no day–night differences in Kcnh3/Kcnh8 transcript expression, exploring this possibility must await the availability of validated anti-Kv12.1 and anti-Kv12.2 antibodies. It is also possible that accessory subunits play a critical role in regulating day–night differences in Kv12-encoded K+ currents densities, as has been demonstrated for the accessory β2 subunit of Kcnma1-encoded BK currents (Whitt et al., 2016).
A rather large number of intracellular and transmembrane accessory subunits of Kv channels have been identified (Paul et al., 2016), and members of at least one subfamily, the Kcne subfamily, have been shown to regulate the expression and properties of Kv12.2-encoded K+ channels (Clancy et al., 2009). Reducing Kcne1 or Kcne3 expression in Xenopus oocytes, for example, increased the amplitudes of heterologously expressed Kv12.2 currents, whereas overexpression of Kcne1 or Kcne3 reduced Kv12.2 current amplitudes (Clancy et al., 2009). Additional experiments revealed that manipulating Kcne1 or Kcne3 affected the surface expression of Kv12.2, whereas the total Kv12.2 protein was not affected (Clancy et al., 2009). Although Kcne1 and Kcne3, as well as other Kcne subfamily members, are expressed in the SCN (http://circadb.hogeneschlab.org), it is not clear whether any of these transcripts and/or the proteins encoded by these transcripts display daily rhythms in expression. It will be important to explore this possibility directly, as well as to determine the functional consequences of manipulating Kcne1 or Kcne3 expression on the repetitive firing properties of SCN neurons.
Posttranslational modifications of the Kv12.1 and/or Kv12.2 α subunits or of accessory subunits of Kv12-encoded channels could also play a role in modulating Kv12 current densities in SCN neurons by influencing cell-surface channel expression directly or by modifying the conductance and/or the open probability of single Kv12 channels. In this context, it is of interest to note that in vivo constitutive activation of the two isoforms of the serine/threonine kinase glycogen synthase kinase 3, GSK3, previously shown to display rhythms in expression and phosphorylation of components of the molecular clock (Iwahana et al., 2004; Iitaka et al., 2005), altered the repetitive firing rates of SCN neurons and disrupted rhythmic locomotor activity (Paul et al., 2012). Subsequent work revealed that GSK3 regulates the amplitude of the persistent component of the Na+ current, INaP (Jackson et al., 2004), in SCN neurons, and that GSK3 inhibition selectively reduced the spontaneous repetitive firing rates of SCN neurons during the day (Paul et al., 2016). It will be of interest to determine if GSK3, or downstream targets of GSK3, plays a role in regulating day–night differences in Kv12-encoded K+ current densities in the SCN.
Physiological implications
The direct link between the spontaneous firing of action potentials and the output of the SCN was established with the in vivo demonstration that continuous infusion of the voltage-gated Na+ channel toxin, TTX, into the SCN of unanesthetized and unrestrained animals (rats) disrupted the circadian rhythm of drinking (Schwartz et al., 1987). Additional in vivo experiments revealed a negative correlation between the repetitive firing rates of SCN neurons and wheel-running activity, i.e., wheel-running activity was high at night when repetitive firing rates were low and, conversely, wheel-running activity was low during the day when repetitive firing rates were high (Earnest et al., 1991). In addition, the acute silencing of SCN neurons during the day (by application of TTX) triggered wheel running, an observation interpreted as demonstrating that repetitive firing rates directly determine the temporal profile of behavioral activity (Earnest et al., 1991). In vitro experiments on SCN explants also showed that the application of TTX eliminated firing and circadian rhythms in vasopressin release (Houben et al., 2014).
Given these combined observations, the finding here that the loss of day–night differences in the repetitive firing rates of SCN neurons with the loss/knockdown of Kv12.1/Kv12.2 did not affect the period of wheel-running activity seems surprising. As noted above, however, previous studies revealed that manipulating Kcnc-encoded FDR (Itri et al., 2005; Kudo et al., 2011) or Kcnma1-encoded BK (Meredith et al., 2006; Montgomery et al., 2013) K+ currents reduced/eliminated day–night differences in repetitive firing rates in the SCN, while having little or no effect on rhythmic wheel-running activity (Itri et al., 2005; Meredith et al., 2006; Kudo et al., 2011; Montgomery et al., 2013). One interpretation of the marked difference in the functional consequences of applying TTX versus changing individual K+ conductances is that the cellular effects are very different. Manipulating individual K+ currents, for example, may alter the daytime and/or nighttime repetitive firing rates of SCN neurons, but, unlike TTX, does not eliminate repetitive firing altogether. In addition, whereas the application of TTX silences all SCN neurons, the impact of manipulating individual K+ currents might be highly variable across cells depending on the repertoire and densities of the K+ and non-K+ channels contributing to shaping individual action potentials and repetitive firing patterns in different cell types (Pennartz et al., 1998; Hermanstyne et al., 2016; Patton et al., 2020). Indeed, the experiments here revealed that the densities of the CX4-sensitive K+ currents were quite variable among nighttime SCN neurons. In addition, ∼30% of nighttime SCN neurons were unaffected by CX4, indicating that the day–night switch in the repetitive firing properties of this subpopulation(s) of neurons is regulated by distinct (non-Kv12-encoded) K+ conductance(s). Given the diversity of cell types in the SCN, distinguished based on morphological features, transmitter phenotypes, synaptic connectivities, and firing properties (Pennartz et al., 1998; Hermanstyne et al., 2016; Allen et al., 2017; Belle and Allen, 2018; Hastings et al., 2018, 2019; Mazuski et al., 2018; Harvey et al., 2020; Michel and Meijer, 2020; Patton et al., 2020), future studies aimed at identifying the critical ionic conductances regulating the intrinsic membrane properties and the day–night oscillations in the repetitive firing rates of SCN should be conducted on identified SCN cell types.
Data availability
Acknowledgments
Olaf S. Andersen served as editor.
The authors wish to thank colleagues in the Nerbonne and Herzog laboratories for helpful discussions and Mr. Richard Wilson for technical assistance. We also thank Dr. Randall Rasmusson of Cytocybernetics for assistance with the development of the IKv12 and IA models and the implementation of these models in dynamic clamp recordings from SCN neurons.
Financial support provided by the National Institute of General Medical Sciences (grant R01 GM104991 to E.D. Herzog and J.M. Nerbonne) and the National Institute of Health (grant R01 NS069842 to TJJ) is gratefully acknowledged. T.O. Hermanstyne was supported in part by a postdoctoral fellowship award from the United Negro College Fund-Merck Science Initiative. All interfering RNAs (RNAi) were obtained from the Washington University RNAi Core, supported by the McDonnell Genome Institute and the Children’s Discovery Institute (grant CDI-LI-2010-94), and all viruses were generated in the Hope Center Viral Vectors Core, supported by a Neuroscience Blueprint Core grant (P30 NS057105).
Author contributions: T.O. Hermanstyne, D.G. Franados-Fuentes, E.D. Herzog, and J.M. Nerbonne designed the research; X. Li and T. Jegla generated the Kv12.1−/− and Kv12.2−/− mouse lines; T.O. Hermanstyne, N.-D. Yang, D. Granados-Fuentes, X. Li, and R.L. Mellor performed the experiments; T.O. Hermanstyne, N.-D. Yang, D. Granados-Fuentes, R.L. Mellor, and J.M. Nerbonne analyzed the data; T.O. Hermanstyne, J.M. Nerbonne, N.-D. Yang, D. Granados-Fuentes, T. Jegla, and E.D. Herzog wrote, commented on, and/or edited the manuscript.
References
This work is part of a special issue on Structure and Function of Ion Channels in Native Cells and Macromolecular Complexes.

