


ADP-ribosylation factor (ARF) GTPases are major regulators of cellular membrane homeostasis. High sequence similarity and multiple, possibly redundant functions of the five human ARFs make investigating their function a challenging task. To shed light on the roles of the different Golgi-localized ARF members in membrane trafficking, we generated CRISPR-Cas9 knockins (KIs) of type I (ARF1 and ARF3) and type II ARFs (ARF4 and ARF5) and mapped their nanoscale localization with stimulated emission depletion (STED) super-resolution microscopy. We find ARF1, ARF4, and ARF5 on segregated nanodomains on the cis-Golgi and ER-Golgi intermediate compartments (ERGIC), revealing distinct roles in COPI recruitment on early secretory membranes. Interestingly, ARF4 and ARF5 define Golgi-tethered ERGIC elements decorated by COPI and devoid of ARF1. Differential localization of ARF1 and ARF4 on peripheral ERGICs suggests the presence of functionally different classes of intermediate compartments that could regulate bi-directional transport between the ER and the Golgi. Furthermore, ARF1 and ARF3 localize to segregated nanodomains on the trans-Golgi network (TGN) and are found on TGN-derived post-Golgi tubules, strengthening the idea of distinct roles in post-Golgi sorting. This work provides the first map of the nanoscale organization of human ARF GTPases on cellular membranes and sets the stage to dissect their numerous cellular roles.
Introduction
The regulation of cellular membrane homeostasis is essential for cell survival. Receptors and important signaling molecules rely on the secretory pathway for effective transport to their correct intracellular destination. ADP-ribosylation factor (ARF) GTPases have emerged as key players in membrane homeostasis and in the regulation of global intracellular trafficking (Adarska et al., 2021). ARFs are a ubiquitous and cross-functional family of small GTPases implicated in many cellular processes from membrane trafficking to metabolism and organization of the cytoskeleton (Jackson and Bouvet, 2014). GDP-GTP exchange factors (GEFs) load ARFs with GTP, triggering conformational changes that allow binding of downstream effectors. GTP hydrolysis mediated by GTPase activating proteins (GAPs) regulates the duration of signal output driven by ARFs (Sztul et al., 2019). Human ARFs are sub-divided into type I (ARF1 and ARF3), type II (ARF4 and ARF5), and type III (ARF6) according to sequence similarity. Type I and type II ARFs share 96 and 90% of their amino acid sequence, respectively, with 80% sequence homology shared between members of the two types. Although different ARFs bind to different subsets of downstream effectors, the switch I and II regions in ARFs, which are responsible for effector binding, are almost identical (Goldberg, 1998; Pasqualato et al., 2001). ARFs’ high sequence similarity has limited the possibility to discern endogenous ARFs in living cells due to the lack of available antibodies, hindering our full understanding of the signaling pathways elicited by the different family members.
ARF1 is the best-known member and its role in the formation of COPI vesicles at the Golgi has been extensively studied using in vitro reconstitution experiments and budding assays from purified Golgi or from Golgi membranes in semi-permeabilized cells (Adolf et al., 2013; Orci et al., 1986; Reinhard et al., 2003). These experiments have led to a great molecular understanding of the ARFs’ on/off cycle. However, ARFs’ functional specificity is driven by activation on a specific membrane and recruitment of numerous effectors, requiring experiments in living cells to fully understand the cellular pathways involved. For example, ARF1, ARF4, and ARF5 have all been shown to support the formation of COPI vesicles in vitro (Popoff et al., 2011). However, it is unclear what happens in the cellular context and why many ARF paralogs are required.
In the last two decades, ARFs have been implicated in different cellular functions beyond their role in Golgi trafficking, for which they were initially discovered. Early studies employing siRNA to deplete ARFs in order to investigate their function showed that type I and type II ARFs have various roles besides Golgi trafficking (Volpicelli-Daley et al., 2005). Type I and II ARFs have all been shown to have a role in TGN export and trafficking in downstream recycling endosomal compartments (Bottanelli et al., 2017; Kondo et al., 2012; Lowery et al., 2013; Manolea et al., 2010; Nakai et al., 2013; Tan et al., 2020). While ARF4 and ARF5 are associated with the ER-Golgi intermediate compartment (ERGIC; Chun et al., 2008), they may additionally have a role in secretion and TGN export in specialized cell types (Deretic et al., 2005; Sadakata et al., 2010).
With multiple and possible redundant functions, studying the role and endogenous distribution of ARFs in living cells has proved to be a challenging task. The observation that ARF pairs needed to be depleted from cells to yield a trafficking defect (Volpicelli-Daley et al., 2005) led to the hypothesis that ARFs may act in pairs. A recent publication showed that GEF-dependent activation of two closely spaced ARF molecules promotes vesicle formation (Brumm et al., 2020). Additionally, ARF dimerization was shown to have a role in vesicle scission (Beck et al., 2011; Beck et al., 2008). Exogenous expression of tagged ARFs, perturbations via GTP-locked mutants, selective siRNA-mediated knockdowns as well as knockouts (KOs) harnessing CRISPR-Cas9 technology (Pennauer et al., 2022) have been the methods utilized so far to reveal specific cellular functions of ARFs. Furthermore, the observation of the intracellular distribution of ARFs has been limited to diffraction-limited microscopy techniques. The Golgi and its neighboring organelles, such as the ERGIC, are tightly packed in the perinuclear area, making super-resolution microscopy essential for investigation of the nanoscale localization of ARFs on different nanodomains or segregating cisternae of the Golgi.
To overcome these limitations, we have generated CRISPR-Cas9 knockin (KI) cell lines of type I (ARF1 and ARF3) and type II ARFs (ARF4 and ARF5) with either the self-labeling enzymes Halo or SNAP for live-cell stimulated emission depletion (STED) microscopy experiments or with ALFA (Götzke et al., 2019) or V5 epitope tags to perform STED microscopy experiments in fixed cells. STED microscopy has allowed us to reveal the nanoscale distribution of each ARF GTPase and to further investigate their intra-Golgi distribution as well as their localization to closely juxtaposed organelles such as the cis-Golgi and ERGIC.
Results
Endogenous tagging of ARF GTPases to investigate their intracellular distribution
To investigate the nanoscale distribution of endogenous ARF GTPases, we generated CRISPR-Cas9 KIs of ARF1, ARF3, ARF4, and ARF5. The only type III ARF member, ARF6, which has a role in endocytosis (Donaldson, 2003), was not included, as this study focuses on Golgi-localized type I and II ARFs. The self-labeling enzyme Halo or the small epitope tag ALFA was introduced at the C-terminus of ARFs to be able to perform STED microscopy experiments.
All endogenously tagged ARFs localized to the Golgi apparatus and to vesicular and tubular-vesicular structures distributed throughout the cytoplasm (Fig. 1). Halo tagging and live-cell microscopy (Fig. 1, A–D) are necessary to visualize ARFs’ tubular-vesicular structures that are otherwise disrupted upon fixation (Fig. 1, E–H). Complementing the live-cell microscopy performed on Halo-tagged ARFs, tagging with 2xALFA allowed significant amplification of the signal of lower abundance endogenous ARFs (like ARF5), thus allowing the investigation of their intra-Golgi localization at higher resolution. Both ARF1EN-Halo and ARF4EN-Halo (Fig. 1, A and C) highlight tubular-vesicular structures, similar to previous observations for ARF1 (Bottanelli et al., 2017). Endogenous ARF3EN-Halo with a glycine/serine (GS) linker between the C-terminus of the protein and the tag displayed strong cytoplasmic localization (Fig. S1). However, the addition of a long flexible linker in the form of a localization and affinity purification (LAP) tag, with eGFP substituted for the Halo tag (Cheeseman and Desai, 2005), revealed the localization of endogenous ARF3 at the Golgi and on tubular-vesicular structures (Fig. 1 B). Interestingly, the presence of either a GS linker or a LAP tag did not affect the localization of other endogenously tagged ARFs (Fig. S1). ARF5EN-Halo is also enriched in the Golgi area, as well as on vesicles, but less prominently on tubular structures (Fig. 1 D). We quantified the relative abundance of all ARFs in HeLa cells and show that the abundance of ARF1 and ARF4 is similarly high, while ARF3 and ARF5 display ∼70 and ∼20% abundance compared to ARF1 and ARF4 (Fig. 1 I). To test the functionality of the fusion proteins, we generated KIs of ARFs in the haploid cell line HAP1 (Fig. S2, A and B) to show that cells do not show any impairment in KDEL receptor (KDELR) recycling (Fig. S2, C and D) and in the recruitment of the COPI and clathrin coats (Fig. S2, E–G), suggesting that the ability to recruit downstream effectors is not affected. Additionally, double KIs of ARF1/ARF4 and ARF4/ARF5 were viable and showed no obvious phenotypic and growth defects (Fig. S2, H and I), while the respective KOs were not viable (Pennauer et al., 2022). With these KIs in hand, we set out to investigate the endogenous localization of type I and II ARFs in living and fixed cells.

Gene editing with CRISPR-Cas9 highlights the endogenous localization of ARF GTPases. (A–H) ARFs were tagged at their endogenous locus with the self-labeling enzyme Halo (A–D) or 2xALFA tag (E–H). An additional LAP tag linker was added in the case of ARF3 (B). Live cells were stained with the Halo substrate JF646-CA (A, C, and D) and JFX650-CA (B). Fixed cells were immunolabeled with an anti-ALFA primary antibody and ATTO647N-conjugated secondaries. (I) Scatter dot plot with mean and SD represents the quantification of the mean fluorescence intensity at the Golgi area measured in the ARFEN-2xALFA KI cells as described in Materials and methods. ARF1 n = 22; ARF3 n = 22; ARF4 n = 24; ARF5 n = 20; n = number of total cells from at least three independent experiments. Yellow arrows highlight structures positive for endogenoulsy tagged ARFs. All images were smoothed with a Gaussian filter and background subtracted as described in Materials and methods. Mean fluo. int. Golgi area (A.U.) = mean fluorescence intensities at the Golgi area in arbitrary units. Scale bars are 10 and 2 µm in the cropped images.
Gene editing with CRISPR-Cas9 highlights the endogenous localization of ARF GTPases. (A–H) ARFs were tagged at their endogenous locus with the self-labeling enzyme Halo (A–D) or 2xALFA tag (E–H). An additional LAP tag linker was added in the case of ARF3 (B). Live cells were stained with the Halo substrate JF646-CA (A, C, and D) and JFX650-CA (B). Fixed cells were immunolabeled with an anti-ALFA primary antibody and ATTO647N-conjugated secondaries. (I) Scatter dot plot with mean and SD represents the quantification of the mean fluorescence intensity at the Golgi area measured in the ARFEN-2xALFA KI cells as described in Materials and methods. ARF1 n = 22; ARF3 n = 22; ARF4 n = 24; ARF5 n = 20; n = number of total cells from at least three independent experiments. Yellow arrows highlight structures positive for endogenoulsy tagged ARFs. All images were smoothed with a Gaussian filter and background subtracted as described in Materials and methods. Mean fluo. int. Golgi area (A.U.) = mean fluorescence intensities at the Golgi area in arbitrary units. Scale bars are 10 and 2 µm in the cropped images.

Addition of a LAP tag greatly improves the localization of endogenously tagged ARF3. Endogenously tagged ARF GTPases either with a GS linker (ARF1EN-Halo and ARF3EN-Halo) or with a LAP tag linker where GFP was switched with Halo tag (ARF1EN-LAP-Halo and ARF3EN-LAP-Halo) labeled with JFX650-CA. While addition of a LAP tag linker improves the membrane and Golgi localization of ARF3, no differences were observed in the localization of ARF1 with or without a LAP tag linker. All images were smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 10 µm.
Addition of a LAP tag greatly improves the localization of endogenously tagged ARF3. Endogenously tagged ARF GTPases either with a GS linker (ARF1EN-Halo and ARF3EN-Halo) or with a LAP tag linker where GFP was switched with Halo tag (ARF1EN-LAP-Halo and ARF3EN-LAP-Halo) labeled with JFX650-CA. While addition of a LAP tag linker improves the membrane and Golgi localization of ARF3, no differences were observed in the localization of ARF1 with or without a LAP tag linker. All images were smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 10 µm.
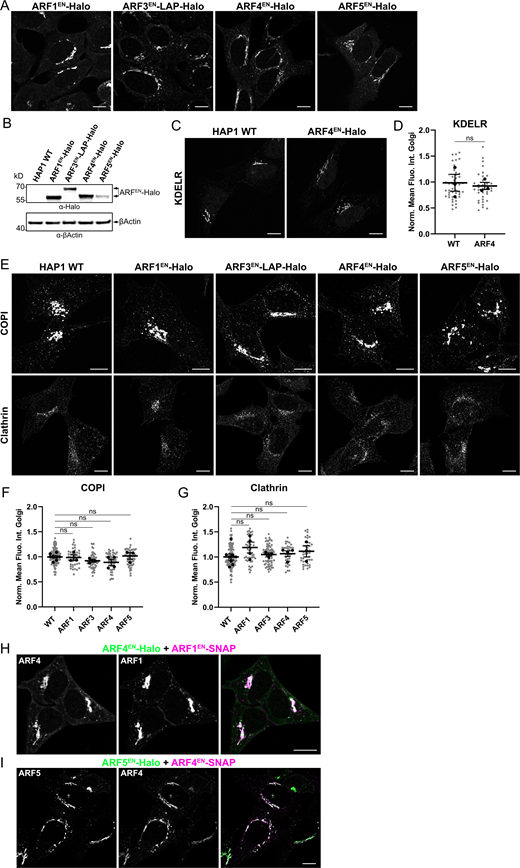
HAP1 cells edited at each ARF locus do not present defects in morphology or coats recruitment. ARFs were tagged at their endogenous locus with the self-labeling enzyme Halo in a HAP1 haploid cell line. (A) Live cells were stained with the Halo substrate JFX650-CA. (B) Immunoblot to detect ARF-Halo fusion proteins in HAP1 KI lysate with anti-Halo antibodies and anti-βActin primary antibody as a loading control. (C and D) HAP1 WT and ARF4EN-Halo KI cells were immunostained with anti-KDELR and secondary antibodies conjugated to Alexa488 (C). Quantification of the normalized mean fluorescence intensity of KDELR at the Golgi area measured in HAP1 WT and ARF4EN-Halo KI cells as described in Materials and methods. Scatter dot plot shows the mean values (black dots) and SEM for each biological replicate (n = 3). All data points (individual cells) are shown as gray dots (n ≥ 10 cells per replicate). Unpaired, two-tailored t test (ns = non-significant, P > 0.05; D). (E–G) HAP1 WT and ARFEN-Halo KI cells were immunostained with anti-COPI and anti-clathrin and secondary antibodies conjugated to Alexa488 (E). Quantification of the mean fluorescence intensity of COPI (F) and clathrin (G) at the Golgi area measured in HAP1 WT and ARFEN-Halo KI cells as described in Materials and methods. Scatter dot plots show the mean values (black dots) and SEM for each biological replicate (n ≥ 3). All data points (individual cells) are shown as gray dots (n ≥ 10 cells per replicate). Ordinary one-way ANOVA versus WT (ns, P > 0.05). (H and I) ARF1EN-SNAP+ARF4EN-Halo (H, magenta/green) and ARF4EN-SNAP+ARF5EN-Halo (I, magenta/green) double KI HAP1 cells were labeled with JF552-CA and JFX650-BG. Norm. Mean Fluo. Int. Golgi = normalized mean fluorescence intensity at Golgi. All images were background subtracted and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 10 µm. Source data are available for this figure: SourceData FS2.
HAP1 cells edited at each ARF locus do not present defects in morphology or coats recruitment. ARFs were tagged at their endogenous locus with the self-labeling enzyme Halo in a HAP1 haploid cell line. (A) Live cells were stained with the Halo substrate JFX650-CA. (B) Immunoblot to detect ARF-Halo fusion proteins in HAP1 KI lysate with anti-Halo antibodies and anti-βActin primary antibody as a loading control. (C and D) HAP1 WT and ARF4EN-Halo KI cells were immunostained with anti-KDELR and secondary antibodies conjugated to Alexa488 (C). Quantification of the normalized mean fluorescence intensity of KDELR at the Golgi area measured in HAP1 WT and ARF4EN-Halo KI cells as described in Materials and methods. Scatter dot plot shows the mean values (black dots) and SEM for each biological replicate (n = 3). All data points (individual cells) are shown as gray dots (n ≥ 10 cells per replicate). Unpaired, two-tailored t test (ns = non-significant, P > 0.05; D). (E–G) HAP1 WT and ARFEN-Halo KI cells were immunostained with anti-COPI and anti-clathrin and secondary antibodies conjugated to Alexa488 (E). Quantification of the mean fluorescence intensity of COPI (F) and clathrin (G) at the Golgi area measured in HAP1 WT and ARFEN-Halo KI cells as described in Materials and methods. Scatter dot plots show the mean values (black dots) and SEM for each biological replicate (n ≥ 3). All data points (individual cells) are shown as gray dots (n ≥ 10 cells per replicate). Ordinary one-way ANOVA versus WT (ns, P > 0.05). (H and I) ARF1EN-SNAP+ARF4EN-Halo (H, magenta/green) and ARF4EN-SNAP+ARF5EN-Halo (I, magenta/green) double KI HAP1 cells were labeled with JF552-CA and JFX650-BG. Norm. Mean Fluo. Int. Golgi = normalized mean fluorescence intensity at Golgi. All images were background subtracted and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 10 µm. Source data are available for this figure: SourceData FS2.
A nanoscale view of the Golgi apparatus with STED super-resolution microscopy
To ultimately pinpoint the localization of ARFs, it was necessary to first obtain a map of the nanoscale localization of known Golgi and vesicular markers with STED microscopy. These included known cisternal markers (GM130, Golgin97, and GRASP65), as well as the vesicular markers COPI and clathrin. The cisternal markers localize to the flat part of the Golgi cisternae, which is tethered to adjacent cisternae, whereas the vesicular markers associate with the highly curved rim of the cisternae where transport intermediates are known to form and bud. STED microscopy allows easy distinction of cis and TGN cisternae, labeled by GM130 and Golgin97, respectively (Fig. 2 A). COPI clusters appeared as puncta around the cis-Golgi cisternae labeled by GM130 (Fig. 2 B, white arrows). STED microscopy also revealed fenestrations in the Golgi cisternae (Fig. 2 B, yellow arrows). COPI clusters appear further away from TGN Golgin97-positive membranes (∼410 nm) in comparison with the cis-Golgi (∼110 nm, Fig. 2 C; and Fig. S3, H and I). COPI-positive clusters represent either forming or fully formed COPI vesicles at the outermost rim of the Golgi. The cisternal markers, GRASP65 and GM130, localize together on the flat/stacked cisternal membranes (Fig. 2 D). Moreover, STED microscopy allows distinction of Golgi-associated ERGIC elements that appear to be tightly tethered to the cis-Golgi cisterna (Fig. 2 E, white arrow). Interestingly, all peripheral (disperse throughout the cytoplasm and away from the perinuclear area) and Golgi-associated ERGICs are decorated by COPI clusters (Fig. 2 F). Clathrin clusters are observed in close proximity to the TGN (∼180 nm, Fig. 2 G and Fig. S3 I) but further apart from the cis-Golgi cisternae (∼530 nm, Fig. 2 H and Fig. S3 H). The same experiments were performed in nocodazole-treated cells where the Golgi ribbon is broken down into smaller Golgi units, known as ministacks (Fig. S3). This approach strongly simplifies the imaging, due to the more favorable geometry, allowing quantification of the distances between cisternae in downstream experiments. The observations made in intact Golgi ribbons are confirmed in nocodazole-treated cells with COPI decorating the rim of cis-Golgi and ERGIC ministacks (Fig. S3, B and C). We show that STED microscopy is instrumental to separate Golgi cisternae and closely juxtaposed membranes, like Golgi-associated ERGICs. We could also easily distinguish nanodomains of clathrin and COPI machinery associated with Golgi cisternae (Fig. 2 I).

STED reveals the super-resolution localization of known Golgi and vesicular markers. (A–H) HeLa cells were immunostained with antibodies against the Golgi markers indicated in the figure and secondaries labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. Line profiles in each panel correspond to the dotted boxes in the cropped images. (B) Yellow arrows highlight fenestrations in the Golgi cisterna, while white arrows highlight COPI-positive vesicular structures. (E) White arrow highlights a Golgi-associated ERGIC structure. (I) Graphical summary of the nanoscale localization of Golgi markers. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
STED reveals the super-resolution localization of known Golgi and vesicular markers. (A–H) HeLa cells were immunostained with antibodies against the Golgi markers indicated in the figure and secondaries labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. Line profiles in each panel correspond to the dotted boxes in the cropped images. (B) Yellow arrows highlight fenestrations in the Golgi cisterna, while white arrows highlight COPI-positive vesicular structures. (E) White arrow highlights a Golgi-associated ERGIC structure. (I) Graphical summary of the nanoscale localization of Golgi markers. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.

Mapping of Golgi markers with STED microscopy in nocodazole-treated cells. (A–G) HeLa cells were treated with nocodazole (33 µM) for 3 h, fixed and immunostained as indicated in the figure. Secondary antibodies labeled either with ATTO647N or AlexaFluor594 were used to be able to perform dual-color STED experiments. (H and I) Scatter dot plots with mean and SD represent the quantification of the distances from the edge of the cis-labeled (H, GM130) or trans-labeled (I, Golgin97) cisternae to the center of COPI and clathrin vesicles measured in the nocodazole-treated fixed cells as described in Materials and methods. GM130: clathrin n = 35; COPI n = 38 (H). Golgin97: clathrin n = 52; COPI n = 56 (I). n = number of ministacks analyzed from at least three independent experiments. All images were background subtracted and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 1 µm.
Mapping of Golgi markers with STED microscopy in nocodazole-treated cells. (A–G) HeLa cells were treated with nocodazole (33 µM) for 3 h, fixed and immunostained as indicated in the figure. Secondary antibodies labeled either with ATTO647N or AlexaFluor594 were used to be able to perform dual-color STED experiments. (H and I) Scatter dot plots with mean and SD represent the quantification of the distances from the edge of the cis-labeled (H, GM130) or trans-labeled (I, Golgin97) cisternae to the center of COPI and clathrin vesicles measured in the nocodazole-treated fixed cells as described in Materials and methods. GM130: clathrin n = 35; COPI n = 38 (H). Golgin97: clathrin n = 52; COPI n = 56 (I). n = number of ministacks analyzed from at least three independent experiments. All images were background subtracted and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 1 µm.
ARF1 localizes to the cis-Golgi while ARF4 and ARF5 localize to Golgi-associated ERGICs
ARF1, ARF4, and ARF5 have all been localized to the ERGIC and cis-Golgi membranes (Chun et al., 2008) and were shown to support COPI vesicle formation in in vitro experiments (Popoff et al., 2011). So why are so many ARFs needed to recruit the COPI coat on early secretory membranes in living cells? COPI has a role in various trafficking steps, including Golgi-to-ER transport and retrograde transport within Golgi cisternae. Additionally, a possible role for COPI in anterograde transport has been suggested and it has also been shown that COPI is associated with Golgi-directed anterograde carriers (Stephens et al., 2000; Weigel et al., 2021). We hypothesize that different ARFs may be recruited to different nanodomains on ERGIC and cis-Golgi membranes to mediate different COPI-dependent trafficking steps.
To test this hypothesis, we mapped ARFs localization on the cis-Golgi and ERGICs with STED microscopy in fixed cells. ARF1 is associated with the cis-Golgi cisternae, labeled by GM130 (Fig. 3, D and G), but excluded from Golgi-associated ERGICs (Fig. 3, A and G). ARF3, the other type I ARF, is never observed on cis-Golgi or ERGIC membranes (Fig. S4). ARF4 and ARF5 decorate the membranes of Golgi-associated and peripheral ERGICs (Fig. 3, B, C, E, F, H, and I). Of the two type II ARFs, ARF4 shows some enrichment on cis-Golgi membranes, albeit at much lower levels than ARF1 (Fig. 3, E and H). Taking advantage of the easier geometry of ministacks in cells that were treated with nocodazole, we were able to quantify the distance between cisternal membranes labeled by the various ARFs and the cis-Golgi marker GM130 (Fig. 3, G–J). While ARF1 localizes to the same cisternae defined by GM130, the average ARF4 and ARF5 signal is mapped ∼400 nm away from the cis-Golgi, confirming their segregated localization to different cisternae (Fig. 3 J). The results are in agreement with observations in intact Golgi ribbons, validating ministacks as a system that greatly simplifies quantification (Tie et al., 2018; Tie et al., 2016).

ARFs segregate on different early secretory membranes. (A–I) ARF1EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were immunostained with anti-ALFA (magenta) and either anti-ERGIC53 (A–C, green) or anti-GM130 (D–F, green) and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. Line profiles in each panel correspond to the dotted boxes in the cropped images. (G–I) ARF1EN-, ARF4EN-, and ARF5EN-2xALFA KI cells were treated with nocodazole (33 µM) for 3 h and immunostained as described above. (J) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae and cis-Golgi cisternae measured in the nocodazole-treated fixed cells as described in Materials and methods. ARF1 n = 9; ARF3 n = 11; ARF4 n = 15; ARF5 n = 14; Golgin97 n = 17. n = number of ministacks from at least three independent experiments. (K) Graphical summary of the nanoscale localization of ARFs on early secretory membranes. Images were deconvolved, background subtracted (A–F), and smoothed with a Gaussian filter (A–I) as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
ARFs segregate on different early secretory membranes. (A–I) ARF1EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were immunostained with anti-ALFA (magenta) and either anti-ERGIC53 (A–C, green) or anti-GM130 (D–F, green) and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. Line profiles in each panel correspond to the dotted boxes in the cropped images. (G–I) ARF1EN-, ARF4EN-, and ARF5EN-2xALFA KI cells were treated with nocodazole (33 µM) for 3 h and immunostained as described above. (J) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae and cis-Golgi cisternae measured in the nocodazole-treated fixed cells as described in Materials and methods. ARF1 n = 9; ARF3 n = 11; ARF4 n = 15; ARF5 n = 14; Golgin97 n = 17. n = number of ministacks from at least three independent experiments. (K) Graphical summary of the nanoscale localization of ARFs on early secretory membranes. Images were deconvolved, background subtracted (A–F), and smoothed with a Gaussian filter (A–I) as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.

STED microscopy reveals ARF3 clusters segregated from ERGIC and cis-Golgi cisternae. (A and B) ARF3EN-2xALFA (magenta) KI HeLa cells were fixed and immunostained with anti-ALFA tag and either anti-ERGIC53 (A, green) or anti-GM130 (B, green). Secondary antibodies labeled with either ATTO647N or AlexaFluor594 were used to be able to perform dual-color STED experiments. (C) ARF3EN-2xALFA KI HeLa cells were treated with nocodazole (33 µM) for 3 h and immunostained with the same primary antibodies described above. Secondary antibodies labeled with either ATTO647N or AlexaFluor594 (GM130)/AlexaFluor568 (ERGIC53) were used. Images were deconvolved, background subtracted (A and B), and smoothed with a Gaussian filter (A–C) as described in Materials and methods. Line profiles in each panel correspond to the dotted boxes in the cropped images. Scale bars are 5 and 1 µm in the cropped images.
STED microscopy reveals ARF3 clusters segregated from ERGIC and cis-Golgi cisternae. (A and B) ARF3EN-2xALFA (magenta) KI HeLa cells were fixed and immunostained with anti-ALFA tag and either anti-ERGIC53 (A, green) or anti-GM130 (B, green). Secondary antibodies labeled with either ATTO647N or AlexaFluor594 were used to be able to perform dual-color STED experiments. (C) ARF3EN-2xALFA KI HeLa cells were treated with nocodazole (33 µM) for 3 h and immunostained with the same primary antibodies described above. Secondary antibodies labeled with either ATTO647N or AlexaFluor594 (GM130)/AlexaFluor568 (ERGIC53) were used. Images were deconvolved, background subtracted (A and B), and smoothed with a Gaussian filter (A–C) as described in Materials and methods. Line profiles in each panel correspond to the dotted boxes in the cropped images. Scale bars are 5 and 1 µm in the cropped images.
When looking at the relative distribution of ARF1, ARF4, and ARF5 in double KI cell lines, ARF1 and ARF4 are partially co-localizing on cis-Golgi membranes, while distinct ARF4-positive Golgi-associated ERGICs devoid of ARF1 are clearly visible (Fig. S5 A). Additionally, ARF4 and ARF5 localize to segregated nanodomains on the ERGIC (Fig. S5 B), while co-localization is observed when ARF1EN-2ALFA cells are labeled with a single anti-ALFA antibody and two different secondary antibodies (Fig. S5 C). These results suggest a functional differentiation between the two type II ARFs. Altogether, these data suggest that ARF1, ARF4, and ARF5 segregate on different early secretory membranes (Fig. 3 K).

STED microscopy of double-edited ARF1-ARF4 and ARF4-ARF5 cells suggests distinct functions. (A and B) ARF1EN-2xV5/ARF4EN-2xALFA (A, green/magenta) and ARF4EN-2xV5/ARF5EN-2xALFA (B, green/magenta) double KI HeLa cells were fixed and immunostained with anti-ALFA and anti-V5 primary antibodies and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. (C) ARF1EN-2xALFA KI HeLa cells were fixed and immunostained with a single anti-ALFA and secondary antibodies labeled with ATTO647N (magenta) and AlexaFluor594 (green). Line profiles in each panel correspond to the dotted boxes in the cropped images. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images (A and B) or 10 and 2 µm in the cropped images (C).
STED microscopy of double-edited ARF1-ARF4 and ARF4-ARF5 cells suggests distinct functions. (A and B) ARF1EN-2xV5/ARF4EN-2xALFA (A, green/magenta) and ARF4EN-2xV5/ARF5EN-2xALFA (B, green/magenta) double KI HeLa cells were fixed and immunostained with anti-ALFA and anti-V5 primary antibodies and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. (C) ARF1EN-2xALFA KI HeLa cells were fixed and immunostained with a single anti-ALFA and secondary antibodies labeled with ATTO647N (magenta) and AlexaFluor594 (green). Line profiles in each panel correspond to the dotted boxes in the cropped images. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images (A and B) or 10 and 2 µm in the cropped images (C).
Live-cell STED highlights segregation of ARF1 and ARF4 on different subclasses of tubular-vesicular early secretory elements
As tubular-vesicular elements at the Golgi and in the cytoplasm are disrupted upon fixation (Fig. 1), we performed live-cell STED experiments to better characterize the distribution of ARFs on Golgi-associated and peripheral tubular-vesicular ERGICs and cis-Golgi-derived structures. For dual-color live-cell STED experiments, Halo and SNAP tag fusions are labeled with cell-permeable Halo chloroalkane (CA) and SNAP benzylguanine (BG) substrates conjugated to STED-compatible organic dyes (Bottanelli et al., 2016; Stockhammer and Bottanelli, 2021). Multi-color live-cell STED microscopy of gene-edited βCOPEN-Halo and transiently expressed SNAP-ERGIC53 highlights COPI clusters on tubular-vesicular ERGIC elements associated with the Golgi ribbon (Fig. S6 A ⅰ) and with peripheral-ERGIC structures further away from the perinuclear area (Fig. S6, A ⅱ and ⅲ). Golgi-associated ERGIC elements are devoid of ARF1EN-Halo and appear as tubular-vesicular structures or cisternae-like structures tethered to the cis-Golgi cisterna labeled by ARF1 (Fig. 4, A ⅰ and ⅱ, white arrows). In addition to defining the cis-Golgi cisternae, ARF1EN-Halo also localizes to a fraction of peripheral ERGICs (Fig. 4 A ⅲ, white arrows), which were not easily detectable in fixed cell imaging. ARF4EN-Halo decorates Golgi-associated ERGICs and a larger subset of peripheral ERGIC53-positive structures (Fig. 4 B). As both ARF4 and ARF1 localize to similar tubular-vesicular structures at the Golgi, we decided to compare the localization of ARF1 and ARF4 in a double KI cell line where ARF4 and ARF1 have been endogenously tagged with Halo and SNAP tag, respectively (Fig. 4 C). ARF4EN-Halo is present on the cis-Golgi cisternae, albeit at a lower level than ARF1, as seen from the partial overlap with the ARF1EN-SNAP signal (Fig. 4 C ⅰ). However, ARF4EN-Halo is strikingly enriched on Golgi-associated ERGICs, which are completely devoid of ARF1EN-SNAP (Fig. 4 C ⅰ, white arrow). Additionally, ARF1EN-SNAP and ARF4EN-Halo define segregated tubular-vesicular elements seen in close proximity to the Golgi (Fig. 4 C ⅱ) or in the cell periphery (Fig. 4 C ⅲ). As ARF1 and ARF4 defined different populations of ERGICs, we quantified the number of peripheral ERGICs positive for these two ARFs in whole cell confocal micrographs (Fig. 4 D and Fig. S6, B–D). While ∼70% of peripheral ERGIC structures are positive for ARF4, only ∼35% are positive for ARF1 and ∼35% are positive for both ARF1 and ARF4 (Fig. 4 D and Fig. S6, B–D). Interestingly, when focusing on the nanoscale localization of ARF1 and ARF4 on peripheral ERGICs with live-cell STED, the two GTPases are seen on segregating nanodomains on average 110 nm apart (Fig. 4 E and Fig. S6 F). To control for chromatic aberrations, ARF1EN-Halo cells were stained with the two different Halo substrates (JF571-CA and JFX650-CA) used for dual-color live-cell STED. The average chromatic shift observed was ∼30 nm (Fig. S6, E and F). These results suggest a differential sorting function for ARF1 and ARF4 at the cis-Golgi–ERGIC interface at the level of sorting tubular-vesicular elements. While ARF1 tubules are retrograde in nature (Bottanelli et al., 2017), ARF4 could contribute to anterograde flow or provide an early recycling platform from the ERGIC to the Golgi (Fig. 4 F).

Live-cell microscopy of tubular-vesicular ERGIC elements and of control cells. (A) βCOPEN-Halo (green) KI HeLa cells were transfected with a plasmid encoding for SNAP-ERGIC53 (magenta). Live cells were stained with the Halo substrate JF571-CA together with the SNAP substrate JFX650-BG to perform dual-color live-cell STED. Yellow arrows highlight COPI-positive clusters associated with tubular-vesicular ERGIC elements highlighted by white arrows. (B and C) ARF1EN-Halo (B, green) and ARF4EN-Halo (C, green) KI HeLa cells were transfected with a plasmid encoding for SNAP-ERGIC53 (magenta) and labeled with the Halo substrate JF552-CA and the SNAP substrate JFX650-BG. White arrows highlight peripheral ERGICs without any ARF, while yellow arrows highlight structures in which ARFs are co-localizing with peripheral ERGICs. (D) ARF4EN-Halo (cyan) and ARF1EN-SNAP (magenta) double KI HeLa cells were transfected with a plasmid encoding for YFP-ERGIC53 (yellow) and labeled with the Halo substrate JF552-CA and the SNAP substrate JFX650-BG. White arrows highlight peripheral ERGICs positive only for ARF4, while yellow arrows highlight peripheral ERGICs positive for both ARF1 and ARF4. (E and F) ARF1EN-Halo KI HeLa cells were stained with the two Halo substrates JF571-CA (green) and JFX650-CA (magenta) used for-dual color live-cell imaging as a control for chromatic aberrations (E). Line profiles in each panel correspond to the dotted boxes in the cropped images. Scatter dot plot with mean and SD represents the quantification of the distances between the two channels in control ARF1EN-Halo cells stained with both dyes (negative control) or between ARF1 and ARF4 nanodomains on peripheral ERGICs (ARF1 vs. ARF4). Examples are shown in Fig. 4 E. ARF1 vs. ARF4 n = 71; negative control n = 56 (F). n = number of ERGICs from at least three independent experiments. Images were deconvolved, background subtracted (A and E), and smoothed with a Gaussian filter (A–E) as described in Materials and methods. Scale bars are: 5 and 1 µm in the cropped images (A); 10 and 2 µm in the cropped images (B–D); 10 µm and 500 nm in the cropped images (E).
Live-cell microscopy of tubular-vesicular ERGIC elements and of control cells. (A) βCOPEN-Halo (green) KI HeLa cells were transfected with a plasmid encoding for SNAP-ERGIC53 (magenta). Live cells were stained with the Halo substrate JF571-CA together with the SNAP substrate JFX650-BG to perform dual-color live-cell STED. Yellow arrows highlight COPI-positive clusters associated with tubular-vesicular ERGIC elements highlighted by white arrows. (B and C) ARF1EN-Halo (B, green) and ARF4EN-Halo (C, green) KI HeLa cells were transfected with a plasmid encoding for SNAP-ERGIC53 (magenta) and labeled with the Halo substrate JF552-CA and the SNAP substrate JFX650-BG. White arrows highlight peripheral ERGICs without any ARF, while yellow arrows highlight structures in which ARFs are co-localizing with peripheral ERGICs. (D) ARF4EN-Halo (cyan) and ARF1EN-SNAP (magenta) double KI HeLa cells were transfected with a plasmid encoding for YFP-ERGIC53 (yellow) and labeled with the Halo substrate JF552-CA and the SNAP substrate JFX650-BG. White arrows highlight peripheral ERGICs positive only for ARF4, while yellow arrows highlight peripheral ERGICs positive for both ARF1 and ARF4. (E and F) ARF1EN-Halo KI HeLa cells were stained with the two Halo substrates JF571-CA (green) and JFX650-CA (magenta) used for-dual color live-cell imaging as a control for chromatic aberrations (E). Line profiles in each panel correspond to the dotted boxes in the cropped images. Scatter dot plot with mean and SD represents the quantification of the distances between the two channels in control ARF1EN-Halo cells stained with both dyes (negative control) or between ARF1 and ARF4 nanodomains on peripheral ERGICs (ARF1 vs. ARF4). Examples are shown in Fig. 4 E. ARF1 vs. ARF4 n = 71; negative control n = 56 (F). n = number of ERGICs from at least three independent experiments. Images were deconvolved, background subtracted (A and E), and smoothed with a Gaussian filter (A–E) as described in Materials and methods. Scale bars are: 5 and 1 µm in the cropped images (A); 10 and 2 µm in the cropped images (B–D); 10 µm and 500 nm in the cropped images (E).

ARF1 and ARF4 define different populations of ERGICs. (A and B) ARF1EN-Halo and ARF4EN-Halo KI HeLa cells were transfected with a plasmid encoding for SNAP-ERGIC53. Live cells were stained with the Halo substrate JF571-CA and the SNAP substrate JFX650-BG and imaged with a STED microscope. (A ⅰ–ⅲ and B ⅰ–ⅲ) White arrows indicate Golgi-associated ERGICs devoid of ARF1 (A i and ii) and positive for ARF4 (B i and ii) and peripheral ERGICs positive for either ARF1 (A iii) or ARF4 (B iii). (C) ARF1EN-SNAP and ARF4EN-Halo double KI HeLa cells were labeled with JF571-CA and JFX650-BG and imaged with a STED microscope. (C ⅰ–ⅲ) Overview of Golgi area showing ARF4 Golgi-associated ERGICs devoid of ARF1 (C i, white arrow) and ARF1/ARF4 segregated tubular-vesicular structures (C ii and iii, yellow and white arrows, respectively). (D) Distribution of peripheral ERGIC populations defined by ARF1 and ARF4. The percentage of peripheral ERGICs positive for either ARF1 or ARF4 was calculated in images of either ARF1EN or ARF4EN-Halo KI cells transfected with a plasmid encoding for SNAP-ERGIC53. The percentage of ARF1- and ARF4-positive peripheral ERGICs was calculated in images of ARF1EN-SNAP and ARF4EN-Halo double KI cells transfected with a plasmid encoding for YFP-ERGIC53. ARF1+ARF4/ERGIC n = 13; ARF1/ERGIC n = 10; ARF4/ERGIC n = 10. n = number of cells from at least three independent experiments. Error bars represent mean and SD. (E) Gallery of peripheral ERGICs showing ARF1/ARF4 segregated nanodomains. (F) Graphical summary of the nanoscale localization of ARF1 and ARF4 on ERGICs. All images were deconvolved, background subtracted, and smoothed as described in Materials and methods. The brightness in the crops was enhanced to highlight the dim peripheral structures. Scale bars are 5 and 1 µm in the cropped images.
ARF1 and ARF4 define different populations of ERGICs. (A and B) ARF1EN-Halo and ARF4EN-Halo KI HeLa cells were transfected with a plasmid encoding for SNAP-ERGIC53. Live cells were stained with the Halo substrate JF571-CA and the SNAP substrate JFX650-BG and imaged with a STED microscope. (A ⅰ–ⅲ and B ⅰ–ⅲ) White arrows indicate Golgi-associated ERGICs devoid of ARF1 (A i and ii) and positive for ARF4 (B i and ii) and peripheral ERGICs positive for either ARF1 (A iii) or ARF4 (B iii). (C) ARF1EN-SNAP and ARF4EN-Halo double KI HeLa cells were labeled with JF571-CA and JFX650-BG and imaged with a STED microscope. (C ⅰ–ⅲ) Overview of Golgi area showing ARF4 Golgi-associated ERGICs devoid of ARF1 (C i, white arrow) and ARF1/ARF4 segregated tubular-vesicular structures (C ii and iii, yellow and white arrows, respectively). (D) Distribution of peripheral ERGIC populations defined by ARF1 and ARF4. The percentage of peripheral ERGICs positive for either ARF1 or ARF4 was calculated in images of either ARF1EN or ARF4EN-Halo KI cells transfected with a plasmid encoding for SNAP-ERGIC53. The percentage of ARF1- and ARF4-positive peripheral ERGICs was calculated in images of ARF1EN-SNAP and ARF4EN-Halo double KI cells transfected with a plasmid encoding for YFP-ERGIC53. ARF1+ARF4/ERGIC n = 13; ARF1/ERGIC n = 10; ARF4/ERGIC n = 10. n = number of cells from at least three independent experiments. Error bars represent mean and SD. (E) Gallery of peripheral ERGICs showing ARF1/ARF4 segregated nanodomains. (F) Graphical summary of the nanoscale localization of ARF1 and ARF4 on ERGICs. All images were deconvolved, background subtracted, and smoothed as described in Materials and methods. The brightness in the crops was enhanced to highlight the dim peripheral structures. Scale bars are 5 and 1 µm in the cropped images.
Different ARF paralogs support COPI recruitment on different ERGIC/Golgi membranes
The differential localization of ARF1, ARF4, and ARF5 to the cis-Golgi and ERGICs indicates that different ARFs may be responsible for COPI recruitment on different early secretory membranes. While all ARFs were shown to recruit COPI in vitro, the localization and proximity of COPI to each ARF paralog has never been explored in living cells. Taking advantage of our gene editing approach combined with live-cell STED, we generated double KIs where ARFs were tagged with a Halo tag, whereas the β subunit of the COPI coat has been tagged with a SNAP tag to investigate the localization of ARF paralogs and COPI machinery (Figs. 5 and S7). COPI coated vesicular structures are seen in close proximity to ARF1EN-Halo at the cis-Golgi (Fig. 5 A ⅰ, yellow arrows). In line with previous work, ARF1EN-Halo defines tubular-vesicular non-ERGIC structures positive for COPI emanating from the Golgi (Fig. 5 A ⅱ, white arrows) and in the cell periphery (Fig. 5 A ⅲ, white arrows). These tubules were shown to carry cargoes like KDELR and to be retrograde in nature (Bottanelli et al., 2017). However, ARF1EN-Halo is rarely enriched in COPI-positive structures, suggesting that upon COPI recruitment on the cis-Golgi, GAP-dependent GTP hydrolysis and ARF release occurs very quickly. Live-cell STED is again instrumental in showing that Golgi-associated ARF4EN-Halo ERGIC tubular-vesicular elements are also decorated with COPI, similar to observations for ARF1 (Fig. 5 B). As for ARF1EN-Halo, ARF4EN-Halo is often observed in proximity to COPI-positive structures (Fig. 5 B ⅱ, yellow arrows). Quantification of the average distance between COPI clusters and the membranes defined by the various ARFs in nocodazole-treated cells shows that type II ARFs are found to be on average more enriched near (5 nm, ARF5) and within (−30 nm, ARF4) COPI clusters than ARF1 (∼100 nm), confirming the initial qualitative observations (Fig. 5, C and D). A differential enrichment of ARF paralogs in COPI structures in living cells hints at a possible differential regulation by GAPs at the cis-Golgi (ARF1) and ERGIC (ARF4 and ARF5).

Live-cell STED reveals segregated ARF1 and ARF4 subpopulations of tubular-vesicular structures defined by COPI machinery. (A and B) ARF1EN/ARF4EN-Halo (green) and βCOPEN-SNAP (magenta) double KI HeLa cells were labeled with JF571-CA and JFX650-BG and imaged with a STED microscope. ARF1 and ARF4 tubular-vesicular structures are highlighted by white arrows and COPI clusters by yellow arrows. (C) Double KI cell lines ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-Halo (magenta) and βCOPEN-SNAP (green) were labeled with JFX650-CA and JF585-BG and subsequently treated with nocodazole (33 µM) for 3 h. (D) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae to the center of COPI vesicles measured in nocodazole-treated live cells as described in Materials and methods. ARF1 n = 62; ARF3 n = 49; ARF4 n = 49; ARF5 n = 48. n = COPI vesicles from at least three independent experiments. All images were deconvolved, background subtracted, and smoothed as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
Live-cell STED reveals segregated ARF1 and ARF4 subpopulations of tubular-vesicular structures defined by COPI machinery. (A and B) ARF1EN/ARF4EN-Halo (green) and βCOPEN-SNAP (magenta) double KI HeLa cells were labeled with JF571-CA and JFX650-BG and imaged with a STED microscope. ARF1 and ARF4 tubular-vesicular structures are highlighted by white arrows and COPI clusters by yellow arrows. (C) Double KI cell lines ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-Halo (magenta) and βCOPEN-SNAP (green) were labeled with JFX650-CA and JF585-BG and subsequently treated with nocodazole (33 µM) for 3 h. (D) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae to the center of COPI vesicles measured in nocodazole-treated live cells as described in Materials and methods. ARF1 n = 62; ARF3 n = 49; ARF4 n = 49; ARF5 n = 48. n = COPI vesicles from at least three independent experiments. All images were deconvolved, background subtracted, and smoothed as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.

COPI-positive clusters are observed near all ARFs. (A–D) Fixed ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were immunostained with anti-ALFA (magenta) and anti-β′COP (CM1, green) and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. Line profiles in each panel correspond to the dotted boxes in the cropped images. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
COPI-positive clusters are observed near all ARFs. (A–D) Fixed ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were immunostained with anti-ALFA (magenta) and anti-β′COP (CM1, green) and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. Line profiles in each panel correspond to the dotted boxes in the cropped images. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
Type I ARFs ARF1 and ARF3 are the sole ARFs localizing to TGN membranes
After establishing the role of different ARFs in sorting at early secretory organelles, we next wanted to test their role in export from the TGN. As all ARFs have been implicated in post-Golgi trafficking (Adarska et al., 2021), we set out to understand which ones localize to the most distal TGN cisternae and to investigate their spatial relationship with the clathrin machinery. Firstly, the localization of the different ARF members endogenously tagged with 2xALFA was tested with the TGN marker Golgin97. To our surprise, only type I ARFs, ARF1 and ARF3, are seen on the TGN cisternae (Fig. 6, A–D). Taking advantage of nocodazole-treated ministacks and their easier geometry, we show that ARF3 is exclusively enriched around TGN membranes, while ARF1 signal is more dispersed, confirming its widespread Golgi localization from cis to trans (Fig. 6, E and F). Interestingly, cisternae positive for type II ARFs (ARF4 and ARF5) are on average ∼700 nm away from TGN membranes defined by Golgin97, similar to the distance observed between GM130 and the TGN (Fig. 6, E and F). While in HeLa cells ARF4 and ARF5 do not associate with the TGN, cell-type specific roles in TGN export may be present, as previously reported (Deretic et al., 2005; Sadakata et al., 2010).

ARF1 and ARF3 are the sole TGN-localized ARF members. (A–E) ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were immunostained with anti-ALFA tag (magenta) and anti-Golgin97 (green) and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. (E) ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were treated with nocodazole (33 µM) for 3 h and immunostained as described above. (F) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae and Golgin97-labeled TGN cisternae measured in nocodazole-treated fixed cells as described in Materials and methods. ARF1 n = 10; ARF3 n = 12; ARF4 n = 10; ARF5 n = 12; GM130 n = 17. n = number of ministacks from at least three independent experiments. Images were deconvolved, background subtracted (A–D), and smoothed with a Gaussian filter (A–E) as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
ARF1 and ARF3 are the sole TGN-localized ARF members. (A–E) ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were immunostained with anti-ALFA tag (magenta) and anti-Golgin97 (green) and secondary antibodies labeled with either ATTO647N or AlexaFluor594 to be able to perform dual-color STED experiments. (E) ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-2xALFA KI HeLa cells were treated with nocodazole (33 µM) for 3 h and immunostained as described above. (F) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae and Golgin97-labeled TGN cisternae measured in nocodazole-treated fixed cells as described in Materials and methods. ARF1 n = 10; ARF3 n = 12; ARF4 n = 10; ARF5 n = 12; GM130 n = 17. n = number of ministacks from at least three independent experiments. Images were deconvolved, background subtracted (A–D), and smoothed with a Gaussian filter (A–E) as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
Live-cell STED shows that ARF1 and ARF3 define segregating nanodomains on the TGN and localize to TGN-derived clathrin-positive tubular-vesicular carriers
An important function of ARFs is the recruitment of clathrin and adaptors to the TGN for enrichment of cargoes and formation of post-Golgi carriers. Transport intermediates, both vesicular and tubular in nature, have been reported (Stalder and Gershlick, 2020). We previously showed that some anterograde cargoes leave the TGN in ARF1-positive tubular-vesicular carriers (Bottanelli et al., 2017). Taking advantage of live-cell STED to preserve sensitive tubular structures, we wanted to test whether any other ARF defines TGN-associated tubular-vesicular elements defined by the clathrin coat. To this end, we generated double KIs of all ARFs (Halo tagged) and clathrin light chain (CLC, SNAP tagged, Fig. 7). As previously shown, ARF1EN-Halo defines TGN-derived tubular-vesicular structures decorated by clathrin (Fig. 7 A). As for ARF1, clathrin clusters are seen adjacent to cisternae defined by highly homologous ARF3EN-LAP-Halo (Fig. 7 B ⅰ, yellow arrows). ARF3EN-LAP-Halo also defines tubular-vesicular structures decorated by clathrin (Fig. 7, B ⅱ and ⅲ, white arrows). Clathrin clusters (associated with the Golgi, endosomes, or the plasma membrane) densely populate the cell cytoplasm, making it hard to define the proximity of ARFs to clathrin in cells with intact Golgi ribbons. To overcome this issue, we again took advantage of nocodazole-induced ministacks and their simpler imaging geometry (Fig. 7 E). Quantification of the distance of clathrin clusters from the edge of the cisternae defined by the various ARFs reveals that ARF4 and ARF5 are the most distal (∼200 nm on average) ARFs from clathrin-positive structures (Fig. 7, E and F). ARF1 is on average ∼100 nm away from a TGN-associated clathrin vesicle, while ARF3 is the most enriched (Fig. 7, E and F).

Type I ARFs define tubular-vesicular clathrin-positive structures on the TGN and the cell periphery. (A–D) ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-Halo (magenta) and SNAP-CLCaEN (green) double KI HeLa cells were labeled with JF571-CA together with JFX650-BG (A and C) or JFX650-CA and JF585-BG (B and D) and imaged with a STED microscope. ARFs tubular-vesicular structures are highlighted by white arrows and clathrin clusters by yellow arrows. (E) Double KI cell lines were labeled with JFX650-CA and JF585-BG and then treated with nocodazole (33 µM) for 3 h. (F) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae to the center of clathrin vesicles measured in nocodazole-treated live cells as described in Materials and methods. ARF1 n = 33; ARF3 n = 30; ARF4 n = 19; ARF5 n = 19. n = clathrin vesicles from at least three independent experiments. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
Type I ARFs define tubular-vesicular clathrin-positive structures on the TGN and the cell periphery. (A–D) ARF1EN-, ARF3EN-, ARF4EN-, and ARF5EN-Halo (magenta) and SNAP-CLCaEN (green) double KI HeLa cells were labeled with JF571-CA together with JFX650-BG (A and C) or JFX650-CA and JF585-BG (B and D) and imaged with a STED microscope. ARFs tubular-vesicular structures are highlighted by white arrows and clathrin clusters by yellow arrows. (E) Double KI cell lines were labeled with JFX650-CA and JF585-BG and then treated with nocodazole (33 µM) for 3 h. (F) Scatter dot plot with mean and SD represents the quantification of the distances from the edge of the ARF-labeled cisternae to the center of clathrin vesicles measured in nocodazole-treated live cells as described in Materials and methods. ARF1 n = 33; ARF3 n = 30; ARF4 n = 19; ARF5 n = 19. n = clathrin vesicles from at least three independent experiments. All images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. Scale bars are 5 and 1 µm in the cropped images.
While ARF1 and ARF3 differ only in seven amino acids at the very N- and C-termini of the proteins, a functional differentiation for the two GTPases has been suggested (Manolea et al., 2010; Volpicelli-Daley et al., 2005). To test the extent of ARF1 and ARF3 co-localization on TGN membranes and post-Golgi trafficking intermediates, we generated double KIs where both, ARF1 and ARF3, were endogenously tagged with Halo and LAP-SNAP tags, respectively. Many areas of the Golgi defined by ARF1 are devoid of ARF3, representing cis-Golgi cisternae (Fig. 8, A ⅰ and ⅱ). Interestingly, while ARF1 localization is more evenly distributed on the cisternae, ARF3 is organized in clusters and on nanodomains devoid of ARF1 (Fig. 8, A ⅰ and ⅱ, white arrows). The two GTPases also co-localized on TGN-derived clathrin-positive tubular trafficking intermediates emanating from the Golgi (Fig. 8 A ⅲ, white arrows) and in the cell periphery (Fig. 8 A ⅳ, white arrows). Although ARF1 is often considered to be the master regulator for clathrin recruitment at the TGN and ARF-dependent post-Golgi sorting, our results point toward an unexplored role of ARF3 in a pathway that segregates from ARF1-mediated trafficking.

ARF1 and ARF3 define segregated nanodomains on the TGN and localize to TGN-derived tubular-vesicular trafficking intermediates. (A) ARF1EN-Halo and ARF3EN-LAP-SNAP double KI HeLa cells were labeled with JF571-CA and JFX650-BG. (A ⅰ–ⅳ) Crops highlight segregation of gene-edited ARF1 and ARF3 on the Golgi (A i and ii) and their localization on TGN-derived (A iii) and peripheral (A iv) tubular-vesicular trafficking intermediates. ARF3 nanodomains are highlighted by white arrows in the crops or a black arrow in the line profile graph (A i and ii). ARF1- and ARF3-positive tubular-vesicular trafficking intermediates are highlighted by white arrows (A iii and iv). (B) Graphical summary of the nanoscale localization of ARF1 and ARF3. Images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. The brightness in the crops was enhanced to highlight the dim distal structures. Scale bars are 5 and 1 µm in the cropped images.
ARF1 and ARF3 define segregated nanodomains on the TGN and localize to TGN-derived tubular-vesicular trafficking intermediates. (A) ARF1EN-Halo and ARF3EN-LAP-SNAP double KI HeLa cells were labeled with JF571-CA and JFX650-BG. (A ⅰ–ⅳ) Crops highlight segregation of gene-edited ARF1 and ARF3 on the Golgi (A i and ii) and their localization on TGN-derived (A iii) and peripheral (A iv) tubular-vesicular trafficking intermediates. ARF3 nanodomains are highlighted by white arrows in the crops or a black arrow in the line profile graph (A i and ii). ARF1- and ARF3-positive tubular-vesicular trafficking intermediates are highlighted by white arrows (A iii and iv). (B) Graphical summary of the nanoscale localization of ARF1 and ARF3. Images were deconvolved, background subtracted, and smoothed with a Gaussian filter as described in Materials and methods. The brightness in the crops was enhanced to highlight the dim distal structures. Scale bars are 5 and 1 µm in the cropped images.
Discussion
In this report, we demonstrate how defining the nanoscale localization of endogenous ARF GTPases has proven to be beneficial to understand their multiple cellular roles. With the help of gene editing and super-resolution microscopy we could finally map ARFs to their intracellular localization in HeLa cells (Fig. 9), necessary for future understanding of how ARFs act in concert with their regulatory GAPs and GEFs to trigger specific cellular responses at specific locations inside living cells. Analysis of the localization of endogenously tagged ARFs highlights functional differences for ARF1, ARF4, and ARF5 at the ER-Golgi interface and ARF1 and ARF3 in post-Golgi trafficking. ARF1 is the only ARF with ubiquitous localization throughout the Golgi stack, suggesting it may be the sole ARF involved in intra-Golgi transport. Even higher resolution microscopy will be required to resolve transport intermediates connecting cisternae and whether they carry only retrograde or also anterograde-directed cargoes (Orci et al., 2000). Localization of ARF1 and ARF3 on partially segregated nanodomains on the TGN indicates a differential role for type I GTPases in post-Golgi sorting. We want to stress the importance of being able to perform super-resolution microscopy experiments in living cells, as many secretory elements like ERGICs and TGN-derived tubular-vesicular structures are disrupted upon fixation, hindering the investigation of trafficking mechanisms.

Model proposing distinct functions of different ARF paralogs in ER-Golgi transport and TGN export.
Model proposing distinct functions of different ARF paralogs in ER-Golgi transport and TGN export.
All type I and II ARFs were shown to support the recruitment of the COPI coat in vitro (Popoff et al., 2011), raising the question as to why multiple ARF paralogs are needed and whether their function is redundant. Endogenous tagging combined with super-resolution STED microscopy reveals that ARF1 and type II ARFs, ARF4 and ARF5, segregate on early secretory membranes (Figs. 3, 4, and 5). ARF1, and to a less extent ARF4, localize to the cis-Golgi cisternae. Interestingly, type II ARFs are enriched on ERGIC tubular-vesicular elements that appear tethered to the cis-Golgi cisternae, completely devoid of ARF1. ARF1, ARF4, and ARF5 also define COPI-positive peripheral ERGIC membranes in the cytoplasm, with a clear fraction of ERGICs, which are positive only for ARF4. Interestingly, STED microscopy highlights that ARF4 and ARF1 localize to segregating nanodomains on ERGICs, whereas the two GTPases are seen colocalizing with confocal microscopy. Our data suggest that ARF1, ARF4, and ARF5 have differential trafficking roles in the early secretory pathway. While ARF1 tubular-vesicular structures forming at the cis-Golgi are loaded with retrograde KDELR cargoes (Bottanelli et al., 2017), we speculate that ARF4/ARF5 ERGICs may have a role in either anterograde ERGIC-to-Golgi transport or ERGIC-to-ER recycling. ARF1 would mediate recycling of ER residents from the cis-Golgi, while early retrieval from the ERGIC would be mediated by ARF4 and ARF5. This model would agree with mass spectrometry analysis showing that COPI vesicles generated with different ARFs have the same content (Adolf et al., 2019). It is unclear whether ARF/COPI are implicated in anterograde ERGIC-to-Golgi transport or simply provide an earlier retrieval platform. However, ARF1 and ARF4 were previously found to both have a role in the formation of ERGIC-derived tubular carriers (Ben-Tekaya et al., 2010) and our data show them co-localizing on segregating ERGIC nanodomains. Taken together, this would suggest that ARF1 only may be responsible for segregating and sorting retrograde cargoes at the ERGIC and cis-Golgi. Interestingly, ARF4 and ARF5 localize to segregating nanodomains on ERGICs, suggesting a functional differentiation within type II ARFs. Further work would be needed to pinpoint the exact role of ARF4 and ARF5 on ERGIC membranes. Another interesting question is whether different COPI isoforms (Moelleken et al., 2007) may be recruited by different ARF paralogs via yet unknown cytosolic adaptors. Scly1 has been proposed to exclusively interact with ARF4 and ARF5 and lead to the recruitment of γ2-COP (Hamlin et al., 2014). Further analysis of the interactome of ARFs will be required to identify possible adaptors specific for other ARFs and the tools presented here will be instrumental to test interactions and super-resolved localization of the possible adaptor-ARF pairs.
Next, we questioned what leads to the differential recruitment of ARF1, ARF4, and ARF5 on early secretory membranes. Only a single ARF GEF is observed on the cis-Golgi/ERGIC membranes, making it unlikely that differential recruitment is driven by the activating GEF (Monetta et al., 2007). A 16 amino acid sequence in the helix α3 of ARF1 has been shown to bind to the SNARE membrin and to be responsible for the recruitment of ARF1 to early Golgi cisternae (Honda et al., 2005). Additionally, p24 family proteins are known to lead to the recruitment of GDP-bound ARF1 (Gommel et al., 2001; Majoul et al., 1998), raising the possibility that different p24 proteins may bind different ARFs at different cellular locations. Structural information shows that, while ARF1 and ARF6 are structurally very similar in the GTP-bound form, they differ in their GDP-bound form (Goldberg, 1998; Pasqualato et al., 2001). This suggests that the highly homologous ARF1, ARF4, and ARF5 could differentiate between a different subset of effectors in their GDP bound forms. Investigation of the proximity of COPI clusters to the various ARFs highlighted that ARF1, ARF4, and ARF5 are differentially enriched in COPI clusters (Fig. 5), with ARF4 and ARF5 being the most enriched. GTP hydrolysis on ARFs and membrane dissociation could occur very quickly after COPI recruitment, as ARF GAP2 and ARF GAP3 are known to bind to the COPI coat and binding enhances GAP activity (Arakel et al., 2019; Weimer et al., 2008). Interestingly, COPI association to the membrane is maintained, despite loss of ARF in the COPI-positive clusters. So far, very little is known about the dynamics of COPI vesicle uncoating in vivo. Our work suggests that ARF1 may dissociate from the membrane earlier than the COPI coat. Additionally, ARF4 and ARF5 are more strongly enriched in COPI clusters than ARF1, suggesting that the regulation of ARF1 at the cis-Golgi and ARF4/ARF5 at the ERGIC may be controlled by different ARF GAPs.
It has also been hypothesized that ARFs act in pairs. ARF1 dimerization has been shown to be important for fission of carriers (Beck et al., 2011; Beck et al., 2008) and dimerization of an ARF GEF in plant cells has been shown to lead to the recruitment and activation of closely positioned ARF pairs (Brumm et al., 2020). Additionally, knockdown of single ARF GTPases failed to yield any phenotype, while knockdown of ARF pairs affected specific trafficking steps (Kondo et al., 2012; Nakai et al., 2013; Volpicelli-Daley et al., 2005). However, CRISPR-Cas9 KO cell lines of ARF1 and ARF4 were recently shown to have an effect on Golgi membrane homeostasis and retrograde Golgi-to-ER transport, while in other double KO cell lines no additional defects were observed (Pennauer et al., 2022). Interestingly, ARF1 and ARF4 could not be deleted simultaneously. Although at a lower level ARF4 is observed on cis-Golgi membranes with ARF1 (Fig. 4 C), raising the possibility that ARF1 and ARF4 may act together to mediate cis-Golgi-to-ER transport and that they could compensate for each other when only one is depleted. ARF4 and ARF5 partially co-localize on the ERGIC membranes, as ARF1 and ARF3 overlap on the TGN. This is a hint that these ARFs may have shared as well as non-overlapping functions. However, the observed overlap may be due to initial association/activation to the same membranes, followed by segregation triggered by association with different subsets of effectors.
Although ARF4 and ARF5 have been implicated in post-Golgi trafficking, imaging of endogenously tagged ARFs shows that only ARF1 and ARF3 are associated with the TGN (Fig. 6). A limitation of this study is that all editing and imaging is carried out in HeLa cells, in which cell-type-specific functions of ARF4 and ARF5 won’t be present. For example, ARF4 has been implicated in trafficking to the cilium (Deretic et al., 2005). However, we cannot reconcile our observation with a function of ARF4 at endosomes (Nakai et al., 2013) as ARF4 exclusively associates with ERGIC membranes. Our editing tools consist of a resistance cassette that allows for selection of successfully edited cells (Stockhammer et al., 2021 Preprint) and therefore will allow to easily edit different cell types to facilitate the discovery of cell-type specific ARF functions. In future studies, it will be particularly important to pinpoint the role of ARFs in polarized cell types and organ-like model systems due to their more complex membrane organization and to the presence of multiple segregating secretory routes.
ARF1 and ARF3 define TGN-derived tubular trafficking intermediates positive for clathrin (Fig. 7). ARF3 differs from ARF1 in only seven amino acids at the very C- and N-termini and the specific localization of ARF3 to the TGN has been mapped to two single C-terminal residues (A174 and K180) conserved across species (Manolea et al., 2010). Previous reports highlighted differential effects on membrane recruitment of ARF1 and ARF3 when ARF GEFs of the brefeldin A–inhibited guanine nucleotide-exchange factors family are depleted (Manolea et al., 2010). Additionally, differential effects on trafficking when ARF1 and ARF3 are knocked down in combination with other ARFs have been observed (Volpicelli-Daley et al., 2005), strongly suggesting non-overlapping roles in TGN export for type I ARFs. Here, using live-cell STED microscopy, we show that gene edited ARF1 and ARF3 localize to segregating nanodomains on the TGN and are seen together on downstream TGN-derived and peripheral tubular trafficking intermediates (Fig. 8). This differential localization strengthens the idea that ARF1 and ARF3 may have differential sorting functions at the TGN, as well as overlapping functions on post-Golgi tubules. Interestingly, gene edited ARF3 with a GC C-terminal linker fails to correctly localize to the Golgi and post-Golgi carriers and shows high cytoplasmic background (Fig. S1). The addition of a long linker in the form of a LAP tag could possibly free the very C terminus of ARF3 for interactions with important uncharacterized membrane recruiting factors.
Lastly, it is interesting to note that the higher numbers of ARF paralogs in mammals compared to yeast seems to be correlated with the more complex membrane organization of mammalian cells. Yeast does not possess ERGICs and has less complex post-Golgi secretory routes. Defining the function of these important classes of transport regulators will have broader implications on our understanding of the organization of intracellular membranes and cellular membrane homeostasis.
Materials and methods
Cell culture
HeLa cells ATCC CCL-2 (ECACC General Collection) and HAP1 cells (Essletzbichler et al., 2014) were grown at 37°C with 5% CO2 in DMEM (Gibco) supplemented with 10% FBS (Corning), 100 U/liter penicillin, and 0.1 g/liter streptomycin (Thermo Fisher Scientific).
Transfection of plasmid DNA in HeLa cells was carried out using FuGENE (Promega). For the DNA transfection, 2 μg of plasmid DNA and 6 μl of FuGENE per well were used. Transfection of plasmid DNA in HAP1 cells was carried out using a NEPA21 electroporation system (Nepa Gene). Approximately 3 million cells were washed twice with Opti-MEM (Gibco) and then resuspended in 90 μl Opti-MEM with 10 µg of DNA in an electroporation cuvette with a 2-mm gap. HAP1 cells were electroporated with a poring pulse of 200 V, 5 ms pulse length, 50 ms pulse interval, 2 pulses, with decay rate of 10% and + polarity; consecutively with a transfer pulse of 20 V, 50 ms pulse length, 50 ms pulse interval, 5 pulses, with a decay rate of 40% and ± polarity.
Nocodazole (Sigma-Aldrich), G418 (Gibco), and puromycin (Gibco) were used at the following concentrations: nocodazole 33 μM; G418 1 mg/ml (HeLa cells) or 3 mg/ml (HAP1 cells); and puromycin 1 μg/ml (HeLa cells).
Preparation of cells for live-cell imaging
For all of the live-cell imaging experiments, cells were seeded on a glass-bottom dish (3.5 cm diameter, No. 1.5 glass; Cellvis) coated with fibronectin (Sigma-Aldrich). Labeling with Halo and SNAP substrates for imaging was carried out for 1 h using 1 μM stocks. After the staining, cells were washed three times with growth medium to get rid of the excess of dye and left for 1 h in an incubator at 37°C and 5% CO2. To break down the Golgi ribbon into ministacks, cells were first labeled and subsequently treated with nocodazole for 3 h. All live-cell imaging was carried out in FluoBrite DMEM (Gibco) supplemented with 10% FBS, 20 mM Hepes (Gibco), and 1× GlutaMAX (Gibco). All live-cell experiments were carried out at 37°C. See Table S1 for the list of dyes used.
Recombinant plasmids for overexpression
To generate the SNAP-ERGIC53 plasmid, a pC4 vector was linearized with EcoRI and BamHI and SNAP and ERGIC53 fragments were inserted as EcoRI-HindIII and HindIII-BamHI fragments, respectively. SNAP was amplified with SNAP-SPs-EcoRI-sense 5′-TACGTGAATTCATGAGGTCTTTGCTAATCTTGGTGCTTTGCTTCCTGCCCCTGGCTGCTCTGGGGAAAGACAAAGACTGCGAAATGAAGCGC-3′ and SNAP-HindIII-anti-sense 5′-ACGATCAAGCTTTCCTGAGCCACCCAGCCCAGGCTTGCCCAGTCTGTGGCCC-3′. The SNAP sense oligo also contains a short sequence encoding for chicken lysozyme signal peptide. The sequence encoding for ERGIC53 was amplified from YFP-ERGIC53 (Hamlin et al., 2014) with ERGIC53-HindIII-sense primer 5′-GATCGTAAGCTTTACTCAGGAGGGGGTGGCGACGGCATGGGAGGAGATGCTGCGGCT-3′ and ERGIC53-BamHI-anti-sense 5′-CAGTACGGATCCTCAGAAGAATTTTTTGGCAGCTGCTTCTTGCTGAGTCCTGTACA-3′.
CRISPR-Cas9 gene editing
The ARF1 genomic locus (gene ID 375) was targeted as previously described (Bottanelli et al., 2017). The ARF3 genomic locus (gene ID 377) was targeted with the guide RNA: 5′- GGTGCTTTGTTAGGGCTGTCTGG-3′ (protospacer adjacent motif sequence is underlined) located after the stop codon in the ARF3 coding sequence. The ARF4 genomic locus (gene ID 378) was targeted with the guide RNA: 5′-TGAAATTGGATATCTAACCAAGG-3′ located after the stop codon in the ARF4 coding sequence. The ARF5 genomic locus (gene ID 381) was targeted with the guide RNA: 5′-TCAAAGCGCTAACCAGCCAGGGG-3′ located after the stop codon in the ARF5 coding sequence. The COPB1 gene (gene ID 1315) was targeted with the guide RNA: 5′-AAATTAACTGTAAAGCTTCAAGG-3′ located after the stop codon in the COPB1 coding sequence. The CLTA gene (gene ID 1211) was targeted using the guide RNA: 5′-ATGGCTGAGCTGGATCCGTTCGG-3′ located after the start codon in the CLTA coding sequence. All guide RNAs were designed using Benchling (https://www.benchling.com) and no guide RNAs have significant off-target matches when checked with the CRISPR design tool. The guide RNAs were cloned into the pSpCas9(BB)-2A-Puro (pX459) V2.0 (plasmid #62988; Addgene; Ran et al., 2013) by annealing oligoes and ligating them into the vector previously linearized with BpiI (Thermo Fisher Scientific). The guide RNA to target the CLTA locus was cloned into the SpCas9 pX330 plasmid (plasmid #42230; Addgene; Cong et al., 2013) vector using the same strategy described above.
The homology repair (HR) plasmid for ARF1-Halo/SNAP was previously described in Bottanelli et al. (2017). The HR plasmid to generate gene edited ARF3-Halo, ARF4-Halo/SNAP, and ARF5-Halo were constructed starting from pTWIST Amp High Copy plasmids (origin: pMB1). The DNA sequence of the left and right homology arms (∼1 kb each) from ARF3 and ARF5 were synthesized by Twist Bioscience with NheI and BamHI sites to insert the desired tag and a GS rich linker (GSGSGSGSGS). In the case of ARF4, only the right homology arm sequence was synthesized by Twist Bioscience with EcoRI, NheI, and BamHI sites for further downstream manipulations. The ARF4 right homology arm was amplified from genomic DNA using the following oligoes: ARF4-Left-Homolgy-Arm-sense 5′-GTCATGAATTCCTGCGTTAGCCAGGAAGGTCTCGATC-3′ and ARF4-Left-Homology-Arm-anti-sense 5′-TCATACGCTAGCTGAGCCGGAACCAGAGCCTGACCCTGATCCACGTTTTGAAAGCTCATTTGACAGCCAGTCAAGTC-3′ and cloned as an EcoRI-NheI fragment. Halo/SNAP tags were introduced into pTWIST ARF3/ARF4/ARF5 HR plasmids as NheI-BamHI fragments. The Halo tag sequence was amplified out of the pHalo-N1 ARF1-Halo HR plasmid (Bottanelli et al., 2017) using the following primers: NheI-Halo-sense 5′-CTGATGCTAGCATGGCAGAAATCGGTACTGGCTTTC-3′ and Halo-BamHI-anti-sense 5′-TACGAGGATCCTCAAGCGTAATCTGGAACATCGTATGGGTAAGCGTAATCTGGAACATCGTATGGGTAGCCGGAAATCTCGAGCGTCGACAGCCAGC-3′. The SNAP tag sequence was amplified out of the pSNAPf plasmid (New England Biolabs) using the following primers: NheI-SNAP-sense 5′-CTGATGCTAGCGACAAAGACTGCGAAATGAAGCGCA-3′ and SNAP-BamHI-anti-sense 5′-TACGAGGATCCTCACTTGTCGTCATCGTCTTTGTAGTCCTTGTCGTCATCGTCTTTGTAGTCACCCAGCCCAGGCTTGCCCAGTCTGTGGCC-3′. The protospacer adjacent motif site in all HR plasmids was mutagenized to prevent cutting by the Cas9.
To facilitate selection of positive recombinants with the drug G418/Geneticin (Gibco), the Neor/Kanr resistance cassette flanked by LoxP sites was introduced into the ARF3/ARF4/ARF5-Halo HR plasmids as a BamHI-BamHI fragment. This allows selection of the positively edited cells with G418 and subsequent excision of the resistance cassette to restore the endogenous locus (Bottanelli et al., 2017; Schmidt et al., 2016). The LoxP-G418r-LoxP fragment was amplified with the following oligoes LoxP-G418r-sense 5′-ATGTCGGATCCATAACTTCGTATAGCATACATTATACGAAGTTATCCTGAGGCGGAAAGAACCAGCTGTGGAATGTGTGTCAGTTAG-3′ and G418r-LoxP-anti-sense 5′- TGCACGGATCCATAACTTCGTATAATGTATGCTATACGAAGTTATTTTATTCTGTCTTTTTATTGCCGTCATAGCGCGGGTT-3′ from a pEGFP-N1 vector (Clontech).
The HR plasmid to generate gene edited ARF3/ARF4/ARF5-2xALFA were generated by linearizing the respective ARF-Halo HR plasmid with NheI and BamHI sites. To generate the ARF1-2xALFA HR plasmid, a new HR plasmid was synthesized (Twist Bioscience) to introduce NheI and BamHI sites and a GS rich linker in the same way as previously designed for ARF3 and ARF5. The sequence encoding for 2xALFA tag was synthesized as a double-strand DNA fragment (Integrated DNA Technologies) and assembled into the linearized NheI-BamHI ARFs HR plasmids via Gibson assembly. In case of ARF1 and ARF3, a new EcoRI site was introduced between the 2xALFA tag and the BamHI site to facilitate further manipulations and allow the excision of the tag or the resistance cassette independently. In case of ARF4 and ARF5, a new HindIII site was introduced between the 2xALFA tag and the BamHI site. The following double-strand DNA fragments were used:
ARF1-NheI-2xALFA-EcoRI-BamHI: 5′-TGGCTGTCCAATCAGCTCCGGAACCAGAAGGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCAGCAGACTGGAGGAGGAGCTGAGAAGAAGACTGACCGAGGGCAGCCGTCTGGAAGAAGAACTGCGTCGTCGTCTGACCGAATGAGAATTCATCGTAGCGGATCCACGCGACCCCCCTCCCTCTCACTCCTCTTG-3′
ARF3-NheI-2xALFA-EcoRI-BamHI: 5′-TGGCTGGCCAATCAGCTCAAAAACAAGAAGGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCAGCAGACTGGAGGAGGAGCTGAGAAGAAGACTGACCGAGGGCAGCCGTCTGGAAGAAGAACTGCGTCGTCGTCTGACCGAATGAGAATTCATCGTAGCGGATCCAAGCGAGACAGCGCTAACAAAGCACCCCAC-3′
ARF4-NheI-2xALFA-HindIII-BamHI: 5′-GACTGGCTGTCAAATGAGCTTTCAAAACGTGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCAGCAGACTGGAGGAGGAGCTGAGAAGAAGACTGACCGAGGGCAGCCGTCTGGAAGAAGAACTGCGTCGTCGTCTGACCGAATGAAAGCTTATCGTAGCGGATCCATGAAATTCGATATCTAACCAACGACATGT-3′
ARF5-NheI-2xALFA-HindIII-BamHI: 5′-GACTGGCTGTCCCACGAGCTGTCAAAGCGCGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCAGCAGACTGGAGGAGGAGCTGAGAAGAAGACTGACCGAGGGCAGCCGTCTGGAAGAAGAACTGCGTCGTCGTCTGACCGAATGAAAGCTTATCGTAGCGGATCCCCAGCGAGGCGCAGGCCCCTGATGCCCGGA-3′
Again, to facilitate selection of positive recombinants with G418, the Neor/Kanr resistance cassette flanked by LoxP sites was introduced into the ARF1/ARF3/ARF4/ARF5-2xALFA HR plasmids as an EcoRI-BamHI (ARF1 and ARF3) or an HindIII-BamHI (ARF4 and ARF5). The (EcoRI/HindIII)-LoxP-G418r-LoxP-BamHI fragments were amplified from a pEGFP-N1 vector with the following oligoes: (EcoRI/HindIII)-LoxP-G418r-sense 5′-ATGTC(GAATTC/AAGCTT)ATAACTTCGTATAGCATACATTATACGAAGTTATCCTGAGGCGGAAAGAACCAGCTGTGGAATGTGTGTCAGTTAG-3′ and G418r-LoxP-BamHI-anti-sense 5′-TGCACGGATCCATAACTTCGTATAATGTATGCTATACGAAGTTATTTTATTCTGTCTTTTTATTGCCGTCATAGCGCGGGTT-3′. The underlined sequence corresponds to the EcoRI/HindIII sites that were used to insert the resistance cassettes into ARF1 and ARF3 / ARF4 and ARF5 HR plasmids, respectively.
The ARF1/ARF4-2xV5 HR plasmids were generated using the same strategy as for the ARFs-2xALFA HR plasmids using Gibson assembly. The following double-strand DNA fragments were inserted into the NheI-BamHI linearized ARF HR plasmids:
ARF1-NheI-2xV5-EcoRI-BamHI: 5′-TGGCTGTCCAATCAGCTCCGGAACCAGAAGGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCGGCAAGCCCATCCCCAACCCCCTGCTGGGCCTGGACAGCACCGGCGGTAAACCAATTCCAAATCCATTGTTGGGTTTGGATTCTACTTGAGAATTCATCGTAGCGGATCCACGCGACCCCCCTCCCTCTCACTCCTCTTG-3′
ARF4-NheI-2xV5-HindIII-BamHI: 5′-GACTGGCTGTCAAATGAGCTTTCAAAACGTGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCGGCAAGCCCATCCCCAACCCCCTGCTGGGCCTGGACAGCACCGGCGGTAAACCAATTCCAAATCCATTGTTGGGTTTGGATTCTACTTGAAAGCTTATCGTAGCGGATCCATGAAATTCGATATCTAACCAACGACATGT-3′
To select for positively gene edited cells, the sequence of the Puromycin resistance cassette (Puromycinr) flanked by LoxP sites was introduced into the ARF1/ARF4-2xV5 HR plasmids linearized with (EcoRI/HindIII)-BamHI. The (EcoRI/HindIII)-LoxP-Puromycinr-LoxP-BamHI fragments were amplified from a HindIII-mutagenized pPUR vector using the following primers: (EcoRI/HindIII)-LoxP-Puror-sense 5′-ATGTC(GAATTC/AAGCTT) ATAACTTCGTATAGCATACATTATACGAAGTTATCTGTGGAATGTGTGTCAGTTAGGGTGTGGAAAGTCCCCAGGCTCCC-3′ and Puror-LoxP-BamHI-anti-sense 5′-TGCACGGATCCATAACTTCGTATAATGTATGCTATACGAAGTTATCAGACATGATAAGATACATTGATGAGTTTGGACAAACCACAAC-3′. The HindIII site in the Puromycinr cassette in the pPUR vector (Clonetech) was mutagenized with the following oligoes: Puror-HindIII-G342C-sense 5′-CTAGGCTTTTGCAAAAACCTTGCATGCCTGCAGGTC-3′ and Puror-HindIII-G342C-anti-sense: 5′-GACCTGCAGGCATGCAAGGTTTTTGCAAAAGCCTAG-3′.
ARF3-LAP-Halo-2xALFA-LoxP-G418r-LoxP HR and ARF3-LAP-SNAP-2xV5-LoxP-Puromycinr-LoxP HR plasmids were generated by linearizing the ARF3-2xALFA-LoxP-G418r-LoxP HR and ARF3-2xV5-LoxP-Puromycinr-LoxP HR with NheI. The LAP-Halo tag and LAP-SNAP tag were synthesized as double-strand DNA fragments and assembled into the respective NheI linearized ARF3-2xALFA-LoxP-G418r-LoxP HR and ARF3-2xV5-LoxP-Puromycinr-LoxP HR plasmids via Gibson assembly.
The following double-strand DNA fragments were inserted into the NheI linearized ARF3 HR plasmids:
LHA-GS-NheI-LAP-Halo-2xALFA: 5′-TGGCTGGCCAATCAGCTCAAAAACAAGAAGGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCATGTACCCAACTTTCTTGTACAAAGTGGTACCGGTCGCCACCGCAAGCGAGAATCTTTATTTTCAGGGCGCCGCCAAATTCAAAGAAACCGCTGCTGCTAAATTCGAACGCCAGCACATGGACAGCGGAGGTGGAGGTTCAAGTGGTCCCTCAGGTTCGTCGAGCCTGGAAGTTCTGTTCCAGGGGCCCCTGTCGTCTAGCGGTCCCTCGGGTTCTATGGCAGAAATCGGTACTGGCTTTCCATTCGACCCCCATTATGTGGAAGTCCTGGGCGAGCGCATGCACTACGTCGATGTTGGTCCGCGCGATGGCACCCCTGTGCTGTTCCTGCACGGTAACCCGACCTCCTCCTACGTGTGGCGCAACATCATCCCGCATGTTGCACCGACCCATCGCTGCATTGCTCCAGACCTGATCGGTATGGGCAAATCCGACAAACCAGACCTGGGTTATTTCTTCGACGACCACGTCCGCTTCATGGATGCCTTCATCGAAGCCCTGGGTCTGGAAGAGGTCGTCCTGGTCATTCACGACTGGGGCTCCGCTCTGGGTTTCCACTGGGCCAAGCGCAATCCAGAGCGCGTCAAAGGTATTGCATTTATGGAGTTCATCCGCCCTATCCCGACCTGGGACGAATGGCCAGAATTTGCCCGCGAGACCTTCCAGGCCTTCCGCACCACCGACGTCGGCCGCAAGCTGATCATCGATCAGAACGTTTTTATCGAGGGTACGCTGCCGATGGGTGTCGTCCGCCCGCTGACTGAAGTCGAGATGGACCATTACCGCGAGCCGTTCCTGAATCCTGTTGACCGCGAGCCACTGTGGCGCTTCCCAAACGAGCTGCCAATCGCCGGTGAGCCAGCGAACATCGTCGCGCTGGTCGAAGAATACATGGACTGGCTGCACCAGTCCCCTGTCCCGAAGCTGCTGTTCTGGGGCACCCCAGGCGTTCTGATCCCACCGGCCGAAGCCGCTCGCCTGGCCAAAAGCCTGCCTAACTGCAAGGCTGTGGACATCGGCCCGGGTCTGAATCTGCTGCAAGAAGACAACCCGGACCTGATCGGCAGCGAGATCGCGCGCTGGCTGTCGACGCTCGAGATTTCCGGCAGCAGACTGGAGGAGGAGCTGAGAAGAAGACTGACCGAGGGCAGCCGTCTG-3′
LHA-GS-NheI-LAP-SNAP-2xV5: 5′-TGGCTGGCCAATCAGCTCAAAAACAAGAAGGGATCAGGGTCAGGCTCTGGTTCCGGCTCAGCTAGCATGTACCCAACTTTCTTGTACAAAGTGGTACCGGTCGCCACCGCAAGCGAGAATCTTTATTTTCAGGGCGCCGCCAAATTCAAAGAAACCGCTGCTGCTAAATTCGAACGCCAGCACATGGACAGCGGAGGTGGAGGTTCAAGTGGTCCCTCAGGTTCGTCGAGCCTGGAAGTTCTGTTCCAGGGGCCCCTGTCGTCTAGCGGTCCCTCGGGTTCTATGGACAAAGACTGCGAAATGAAGCGCACCACCCTGGATAGCCCTCTGGGCAAGCTGGAACTGTCTGGGTGCGAACAGGGCCTGCACCGTATCATCTTCCTGGGCAAAGGAACATCTGCCGCCGACGCCGTGGAAGTGCCTGCCCCAGCCGCCGTGCTGGGCGGACCAGAGCCACTGATGCAGGCCACCGCCTGGCTCAACGCCTACTTTCACCAGCCTGAGGCCATCGAGGAGTTCCCTGTGCCAGCCCTGCACCACCCAGTGTTCCAGCAGGAGAGCTTTACCCGCCAGGTGCTGTGGAAACTGCTGAAAGTGGTGAAGTTCGGAGAGGTCATCAGCTACAGCCACCTGGCCGCCCTGGCCGGCAATCCCGCCGCCACCGCCGCCGTGAAAACCGCCCTGAGCGGAAATCCCGTGCCCATTCTGATCCCATGTCATCGAGTAGTGCAAGGAGATCTGGACGTGGGGGGCTACGAGGGCGGGCTCGCCGTGAAAGAGTGGCTGCTGGCCCACGAGGGCCACAGACTGGGCAAGCCTGGGCTGGGTGGCAAGCCCATCCCCAACCCCCTGCTGGGCCTGGACAGCACCGGCGGTAAACCAATTCCAAATC-3′
The ARF1-LAP-Halo-PolyA-G418r HR plasmid was generated by linearizing the ARF1-Halo-PolyA-G418r HR plasmid with BamHI. The LAP-Halo tag was synthesized as a double-strand DNA fragment and assembled into the BamHI linearized ARF1-Halo-PolyA-G418r HR plasmid via Gibson assembly.
Sequence of the double-strand DNA fragment inserted:
LHA-GS-BamHI-LAP-Halo: 5′-CTACGAAACCAGAAGGGATCGTCAGGTCGGGATCCAATGTACCCAACTTTCTTGTACAAAGTGGTACCGGTCGCCACCGCAAGCGAGAATCTTTATTTTCAGGGCGCCGCCAAATTCAAAGAAACCGCTGCTGCTAAATTCGAACGCCAGCACATGGACAGCGGAGGTGGAGGTTCAAGTGGTCCCTCAGGTTCGTCGAGCCTGGAAGTTCTGTTCCAGGGGCCCCTGTCGTCTAGCGGTCCCTCGGGTTCTATGGGCTCAGGTTCTGGAATGGCAGAAATCGGTACTGGCTTTC-3′
To generate the SNAP-CLC HR plasmid, SNAP-Sec61Beta (Bottanelli et al., 2016) was digested with BglII and MluI and the right homology arm (∼1 kb) was synthesized by IDT Technologies as a double-strand DNA fragment and assembled into the linearized vector. The resulting vector was then digested with AseI and NheI and the left homology arm (∼1 kb) was assembled using the same strategy. Similar to previously generated constructs, the Puromycinr resistance cassette flanked by LoxP sites was introduced into the SNAP-CLC HR plasmids as a NheI-NheI fragment. The LoxP-Puromycinr-LoxP fragment was amplified from the pPUR vector with the following oligoes: NheI-LoxP-Puror-sense 5′-ATGTCGCTAGCATAACTTCGTATAGCATACATTATACGAAGTTATCTGTGGAATGTGTGTCAGTTAGGGTGTGGAAAGTCCCCAGGCTCC-3′ and Puror-LoxP-NheI-anti-sense 5′- TGCACGCTAGCATAACTTCGTATAATGTATGCTATACGAAGTTATCAGACATGATAAGATACATTGATGAGTTTGGACAAACCACAAC-3′.
To generate the BetaCOP-SNAP/Halo HR plasmid, the sequence containing the left and right homology arms (∼1 kb each) to target the COPB1 locus were synthesized by Thermo Fisher Scientific with NheI and BamHI sites to insert the desired tag and a GS rich linker (GSGSGSGSGS). The SNAP- or Halo-tag was cloned into the linearized plasmid as a NheI-BamHI fragment as previously described for the ARFs-SNAP/Halo HR plasmids. To select for positive clones with G418, the G418 resistance cassette was introduced as a BamHI-PolyA-G418r-BamHI fragment. The resistance cassette was amplified from the ARF1-Halo HR plasmid using the following primers: BamHI-PolyA-sense 5′-ATGTCGGATCCCCGCGACTCTAGATCATAATCAGC-3′ and G418r-BamHI-anti-sense 5′-TCATGGGATCCTTTATTCTGTCTTTTTATTGCCGTC-3′.
HR plasmids and their respective guide RNAs plasmids were introduced into HeLa CCL-2 cells using FuGENE (Promega). G418 (1 mg/ml) and puromycin (1 µg/ml) were added to the cells 3 d after transfection. After selection and recovery, cells were either prepared for downstream applications or they were again transfected with 2 µg of pBS598 EF1alpha-EGFPcre plasmid (plasmid #11923; Addgene) encoding for cre recombinase to excise the resistance cassette, in case a resistance cassette flanked by LoxP was used.
To obtain single cell clones of the ARF3-, ARF4-, and ARF5-Halo KIs, cells were subjected to FACS and single cells were deposited in a well of a 96-well plate. Clones were genotyped via Western blot using anti-Halo and anti-ARF antibodies, fluorescent microscopy, and PCR using the following oligoes: ARF3-genotyping-sense 5′-GGTACATTCAGGCCACCTGTGCCACCAGCGGGGACGGG-3′, ARF3-genotyping-anti-sense 5′-CTCCCACCTTCTTCCCTTAATCTCACCAACACGGAAGGGGC-3′, ARF4-sense 5′-TGTGAGATGCAAGAGGTGTG-3′, ARF4-genotyping-anti-sense 5′-CCAGCCAGAGAAAGATCCAAAACAC-3′, ARF5-sense 5′-TTCCCCACAGTGGTATGTCCA-3′ and ARF5-genotyping-anti-sense 5′-ATACCCTCAGCTCAGCTTCTCCCTC-3′. In case of ARF3, the genotyping PCR did not yield results and the selection of the ARF3 clone (clone 19) for the experiments was based on the fluorescent intensity taken out of the microscopy images (brightest clones) In case of ARF4 and ARF5, homozygous clones could be selected, based on the genotyping PCR results (ARF4: clone 7 and ARF5: clone 14).
For the generation of KI cell lines in HAP1 cells, the ARF1/3/4/5-Halo HR plasmid and their respective guide RNAs plasmids were transfected using a NEPA21 electroporation system (3 × 106 cells, mixed with 5 µg of guide RNA plasmid and 5 µg of HR plasmid). Again, G418 (3 mg/ml) was added 3 d after transfection. After selection and recovery, ARF3/4/5-Halo edited cells were electroporated with 10 µg of pBS598 EF1alpha-EGFPcre to excise the resistance cassette. ARFs-Halo HAP1 cells were also sorted based on cell size and fluorescence to obtain a homogenous population of edited haploid cells (Beigl et al., 2020).
The ARF1EN-Halo and BetaCOPEN-SNAP double KI cell line was generated in Bottanelli et al. (2017). ARF3/ARF4/ARF5-Halo and BetaCOP-SNAP double KI were generated by transfecting BetaCOP-SNAP-polyA-G418 and BetaCOP guide in the sorted ARFs-Halo KIs.
The ARF1/3/4/5EN-Halo and SNAP-CLCaEN double KI cell lines were generated by transfecting LoxP-Puromycinr-LoxP-SNAP-CLCa HR plasmid and CLCa guide into the sorted ARFs-Halo KIs. Similarly, the ARF3EN-LAP-Halo and SNAP-CLCaEN double KI cell line was generated by transfecting the CLCa HR and guide plasmids mentioned above into the ARF3EN-LAP-Halo cell line.
To generate the ARF1-SNAP and ARF4-Halo double KIs, ARF4-Halo KI was transfected with the ARF1-SNAP-LoxP-G418r-LoxP plasmid and the ARF1 guide. The ARF1EN-Halo and ARF3EN-LAP-SNAP double KI cell line was generated by transfecting the respective ARF3 reagents into the ARF1EN-Halo cell line.
The ARF1/ARF3/ARF4/ARF5-2xALFA cell lines were generated by transfecting the HR plasmids and guides into HeLa cells.
To generate the ARF4-2xALFA and ARF1-2xV5 double KI cell line, the ARF4-2xALFA-loxP-Puro-LoxP HR plasmid was transfected with the ARF4 guide into an already existing ARF1-2xV5 cell line. Similarly, the ARF4-2xV5 and ARF5-2xALFA double KI cell line was generated by transfecting the ARF4-2xV5-loxP-Puro-LoxP HR plasmid with the guide into the already existing ARF5-2xALFA cell line.
All the cell lines generated in this study were validated via Western Blot analysis and PCR amplification and sequencing of the edited locus after antibiotic selection and/or excision of the resistance cassette.
Immunofluorescence (IF)
IF was carried out on cells seeded on 1.5# glass 12 mm coverslips previously coated with fibronectin. All steps were carried out at RT. First, samples were rinsed three times with PBS and then fixed using 4% PFA (16% stock, Electron Microscopy Sciences) for 10 min. After fixation, samples were rinsed three times with PBS and then permeabilized using permeabilization buffer (0.3% NP40, 0.05% Triton X-100, and 0.1% BSA [IgG free] in PBS). Cells were rinsed three times with wash buffer (0.05% NP40, 0.05% Triton X-100, and 0.2% BSA in PBS) and blocked for 45 min in block buffer (0.05% NP40, 0.05% Triton X-100, and 5% goat serum [Jackson ImmunoResearch] in PBS). Subsequently, cells were incubated with primary antibodies (1:1,000) for 1 h in block buffer and then washed three times for 5 min with wash buffer. Secondary antibody (1:2,000) incubation was carried out for 30 min in block buffer. Again, samples were washed three times for 5 min with wash buffer and then rinsed three times with PBS. Samples were then dipped in ddH2O and mounted with pro-long Gold (Life Technologies). See Table S2 for the list of antibodies used.
STED imaging
STED imaging was performed on a commercial expert line Abberior STED microscope equipped with 485, 561, and 645 nm excitation lasers and an Olympus Objective UPlanSApo 100× 1.40 NA. The 775 nm depletion laser was used to deplete dyes in two-color STED experiments. Two-color images were recorded sequentially line by line. Detection was carried out with avalanche photodiodes with detection windows set to 571–630 nm and 650–756 nm. When required, excitation power was kept constant between samples to allow the quantification of signal differences. Acquisition was carried out with Instruments Development Team, Imspector Image Acquisition & Analysis Software v16.3.16118 (http://www.imspector.de). The pixel size was set to 60 nm for confocal imaging, 30 nm for live-cell STED imaging, and 20 nm for STED imaging in fixed cells. Live-cell STED imaging was carried out at 37°C.
Image processing and statistical analysis
Raw STED images were deconvolved to reduce noise using Richardson–Lucy deconvolution from the python microscopy PYME package (https://python-microscopy.org/), background subtracted and smoothed using a Gaussian filter with 1-pixel SD using ImageJ (Abramoff et al., 2004) as indicated in the figures.
Mean fluorescence intensities at the Golgi (Fig. 1 I) were measured using ImageJ. To select the area for quantification of the signal intensity at the Golgi, a mask of the Golgi was determined by a thresholding method from ImageJ (MaxEntropy).
Line profile data shown in Fig. 2, 3, 4, 6, and 8 and Fig. S4, S5, and S6 were obtained from Gaussian blurred images using ImageJ by drawing a line perpendicular to the direction of the cisternae. Line profile data from ImageJ were plotted using GraphPad Prism (GraphPad Software, https://www.graphpad.com).
The quantification of ARF1- and ARF4-positive peripheral ERGICs (Fig. 4 D) was performed using raw image data in ImageJ and plotted using GraphPad Prism. Representative images are presented in Fig. S6, B–D. The percentage of peripheral ERGIC positive for either ARF1 or ARF4 was calculated by counting all ARF1/ARF4-positive peripheral ERGICs divided by the total amount of peripheral ERGICs in ARF1EN- and ARF4EN-Halo KIs cells expressing SNAP-ERGIC53. The percentage of ARF1- and ARF4-positive peripheral ERGIC structures was calculated by counting all ARF1- and ARF4-positive peripheral ERGICs divided by the total amount of peripheral ERGICs in ARF1EN-SNAP and ARF4EN-Halo double KI HeLa cells expressing YFP-ERGIC53.
To quantify the distance from the edge of the ARF-labeled cisternae to the GM130-labeled cis-Golgi cisternae (Fig. 3 J) and to the Golgin97-labeled TGN cisternae (Fig. 6 F) in nocodazole-treated fixed ARFsEN-2xALFA KI cells, side-view images of ministacks were chosen. A box perpendicular to the direction of the cisternae was drawn to obtain a line profile using ImageJ. The box profile data were further analyzed with a custom Python code (available on https://github.com/AG-Bottanelli/distance-to-edge.git) that defined the edge of the cisternae. The edge of each cisterna (ARFs, GM130, and Golgin97) was calculated as the coordinate with a maximum mean intensity gradient. After the first computation attempt, we plotted the line profiles along with their gradients and discarded those in which the edge was detected in a second peak of signal for M-shaped profiles. The data obtained from the Python script were plotted using GraphPad Prism.
To calculate the distance from the edge of the ARF-labeled cisternae to the center of COPI-positive (Fig. 5 D) or clathrin-positive (Fig. 7 F) clusters in live nocodazole-treated ARFsEN-Halo+BetaCOPEN-SNAP/SNAP-CLCaEN double KI cells, en face view images of ministacks were chosen and line profiles were obtained by drawing a box between the ARF cisternae and the vesicle using ImageJ. The same approach was used to calculate the distance from the edge of the GM130-labeled (Fig. S3 H) or Golgin97-labeled (Fig. S3 I) cisternae to the center of COPI or clathrin vesicles in nocodazole-treated fixed HeLa cells. Again, the line profile data were further analyzed with a Python script (available on https://github.com/AG-Bottanelli/distance-to-edge.git) that defined the center of a COPI/clathrin cluster and the edge of the cisternae as described previously. The center of COPI/clathrin clusters was determined as the position of the maximum mean signal intensity value. After the first computation attempt, we plotted the line profiles along with their gradients and discarded those in which (1) “vesicle” signal’s full width at half maximum was higher than 200 nm or (2) edge was detected in a second peak of signal for M-shaped profiles. The data obtained from the Python script were plotted using GraphPad Prism.
The quantification of the mean KDELR, COPI, and clathrin signal at the Golgi area in HAP1 cells (Fig. S2, D, F, and G) was measured using raw data in ImageJ. To select the area for quantification of the signal intensity at the Golgi, a mask of the Golgi was determined by using the thresholding function (Moments) from ImageJ. HAP1 WT and HAPI ARFsEN-Halo KI IF samples were prepared and imaged on the same day using the same parameters on the STED microscope to ensure comparability between samples. Signal intensities for quantification come from three independent experiments. The signal intensities from the KDELR/COPI/clathrin signal in the ARFsEN-Halo KI cells were normalized to the mean signal intensity from the WT cells. SuperPlots in Fig. S2 were generated as in Lord et al. (2020) using GraphPad Prism. Unpaired t test (Fig. S2 D) and ordinary one-way ANOVA (Fig. S2, F and G) were carried out with GraphPad Prism and P values are indicated in the figure legend.
To calculate the distance between ARF1- and ARF4-positive peripheral ERGICs (Fig. S6 F and Fig. 4 E), line profiles were obtained by drawing a box perpendicular to the ARF vesicular structure using ImageJ. Afterward, the distance between the maxima from the line profiles of ARF4EN-Halo vs ARF1EN-SNAP was calculated using a custom-made Python script (https://github.com/AG-Bottanelli/distance-to-edge.git) and then plotted using GraphPad Prism. As a negative control, ARF1EN-Halo cells were stained with the Halo substrates JF571-CA and JFX650-CA and as described above; line profiles were obtained by drawing a box perpendicular to the JF571-labeled ARF1 vesicular structure and the same JFX650-labeled ARF1 vesicular structure using ImageJ. Again, the distance between the maxima from the line profiles of ARF1EN-Halo JF571-CA vs. ARF1EN-Halo JFX-650-CA was calculated in an identical way and then plotted using GraphPad Prism (Fig. S6, E and F).
SDS-PAGE and Western blot
Protein lysates of HAP1 and HeLa cells were separated by SDS-PAGE using 4–12% gradient precast polyacrylamide gels (Life Technologies). The proteins separated by SDS-PAGE were transferred onto a nitrocellulose membrane using a Mini-Protean Tetra Cell device. Halo tag fusions were detected using a primary anti-Halo antibody at a dilution of 1:1,000 in PBS supplemented with 1% BSA and 0.02% sodium azide. For the detection of primary antibodies, anti-mouse horseradish peroxidase–coupled (1:5,000) secondary antibodies were used. For final detection, Pierce ECL substrate was used, and bioluminescent signal was detected.
Online supplemental material
Fig. S1 shows that the addition of a LAP tag linker improves the membrane and Golgi localization of endogenously tagged ARF3. Fig. S2 shows that endogenously tagged ARFs in the haploid cell line HAP1 do not present defects in morphology, KDELR trafficking and are able to recruit downstream coats (COPI and clathrin). Fig. S3 displays the spatial relation between Golgi markers with STED microscopy in nocodazole-treated cells. Fig. S4 shows with STED microscopy that endogenous ARF3 is excluded from early secretory compartments (ERGIC and cis-Golgi). Fig. S5 shows STED microscopy images of double-edited ARF1-ARF4 and ARF4-ARF5 cells localizing to segregating nanodomains. Fig. S6 shows live-cell microscopy images of tubular-vesicular ERGIC elements. Fig. S7 shows STED microscopy images of endogenous ALFA-tagged ARFs and COPI in fixed cells. Table S1 lists dyes used for live-cell imaging. Table S2 lists antibodies used in this study.
Data availability
Data presented in this publication are available from the authors upon reasonable request. Images of uncropped blots are available as source data figures.
Acknowledgments
We would like to thank the whole lab for the many discussions and for reading this manuscript. A particular thank you goes to Blanche Schwappach and Andreas Ernst for critically reading the manuscript and for the many engaging discussions on the topic. We thank Jez Carlton for suggesting the use of the LAP tag. We thank Luke Lavis, Jonathan Grimm, and the team of chemists at Janelia (Janelia Research Campus, Ashburn, VA, USA) that provide us with the latest and brightest STEDable dyes for our experiments.
This project was supported by Deutsche Forschungsgemeinschaft (German Research Foundation) grants SFB958 (Project A25 [C. Rodilla-Ramirez and A. Stockhammer]), SFB/TRR186 (Project A20 [L. Wong-Dilworth and P. Adarska]), and a major research instrumentation grant for the acquisition of the STED microscope.
Author contributions: F. Bottanelli and L. Wong-Dilworth conceived the project. L. Wong-Dilworth, C. Rodilla-Ramirez, E. Fox, S.D. Restel, A. Stockhammer, P. Adarska, and F. Bottanelli carried out experiments. C. Rodilla-Ramirez developed the quantification tools. L. Wong-Dilworth and C. Rodilla-Ramirez carried out imaging quantification. A. Stockhammer provided STED expertise. F. Bottanelli and L. Wong-Dilworth wrote the manuscript and all authors reviewed and edited the manuscript.

