Intracellular transport among organellar compartments occurs in two general ways: by membrane-bound carriers and membrane contacts. Specific circumstances that involve the coordination of these two modes of transport remain to be defined. By studying coat protein I (COPI) transport, we find that phosphatidylcholine with short acyl chains (sPC) is delivered through membrane contact from the endoplasmic reticulum (ER) to sites of COPI vesicle formation at the Golgi to support the fission stage. Phosphatidylinositol transfer protein beta (PITPβ) plays a key role in this process, with the elucidation of this role shedding new insights into how PITPβ acts, providing a mechanistic understanding of a specific circumstance when vesicular transport requires membrane contact and contributing to the general understanding of how intracellular transport carriers are formed.

Introduction
Transport by membrane-bound carriers occurs through conserved steps that are mediated by major classes of effectors. Coat complexes initiate transport by bending the membrane to generate carriers and binding to cargoes for their sorting into these carriers. Subsequently, tether complexes act by docking the transport carriers to their target compartments. Soluble NEM-sensitive associated protein (SNAP) receptors (SNAREs) then complete transport by promoting the fusion of carriers with their target compartments. A detailed understanding of how these core transport effectors act and how they are regulated is being achieved for model pathways (Brocker et al., 2010; Cai et al., 2007; Donaldson and Jackson, 2011; McMahon and Boucrot, 2011; Miller and Barlowe, 2010; Mizuno-Yamasaki et al., 2012; Pucadyil and Schmid, 2009; Wickner and Schekman, 2008).
How transport occurs through membrane contacts is also being elucidated. This mode of transport has been found to play widespread roles in the delivery of lipids among organellar compartments. Multiple classes of proteins have been identified to possess lipid transfer activity. These lipid transfer proteins can also participate in contact formation. How these two functions are integrated to achieve lipid transport through membrane contact is also being elucidated (Antonny et al., 2018; Cohen et al., 2018; Lujan et al., 2021; Prinz et al., 2020; Reinisch and Prinz, 2021; Saheki and De Camilli, 2017; Venditti et al., 2020; Wong et al., 2019; Wu et al., 2018).
The ER forms contact with multiple organellar compartments through vesicle-associated membrane protein (VAMP)-associated proteins (VAPs) that reside on the ER membrane (Murphy and Levine, 2016). In the case of cholesterol transfer, VAP-A has been found to interact with oxysterol binding protein (OSBP) on the Golgi membrane to form contact between the ER and the trans-Golgi network (TGN). OSBP also possesses sterol transfer activity, and this activity is promoted by the countertransfer of phosphatidylinositol 4-phosphate (PI4P) by OSBP. Through reconstitution studies, a mechanistic understanding of how OSBP couples its functions in lipid transfer and contact formation is being achieved (Mesmin et al., 2013).
Studies on vesicle formation by the coat protein I (COPI) complex have been contributing to a general understanding of how transport carriers are generated (Hsu et al., 2009; Jackson, 2014; Lippincott-Schwartz and Liu, 2006; Pucadyil and Schmid, 2009). COPI vesicles act in retrograde transport at the Golgi complex and also from the Golgi to the endoplasmic reticulum (ER) (Brandizzi and Barlowe, 2013; Lee et al., 2004; Suda et al., 2018). Early studies identified coatomer, a multimeric complex, as the core component of the COPI complex, and ADP-ribosylation factor 1 (ARF1) as the small GTPase that regulates the recruitment of coatomer from the cytosol to the Golgi membrane to initiate COPI vesicle formation (Donaldson et al., 1992; Malhotra et al., 1989; Serafini et al., 1991; Waters et al., 1992). Subsequently, a GTPase activating protein (GAP) that deactivates ARF1 (known as ARFGAP1) was identified to act not only as its regulator (Cukierman et al., 1995) but also as an effector (Yang et al., 2002) in COPI vesicle formation.
The late stage of vesicle formation involves membrane fission, which severs the neck of coated buds to release them as coated vesicles. Brefeldin-A ADP-ribosylation substrate (BARS) has been identified to promote COPI vesicle fission (Yang et al., 2005). More recently, lipid-based mechanisms have also been discovered to play a key role (Park et al., 2019; Yang et al., 2008, 2011). COPI vesicle fission has been found to require the sequential actions of phospholipase D2 (PLD2) activity, which converts phosphatidylcholine (PC) to phosphatidic acid (PA), followed by lipid phosphate phosphatase 3 (LPP3) activity, which converts PA to diacylglycerol (DAG) (Park et al., 2019). The critical roles of PA and DAG require these lipids to have short acyl chains (Park et al., 2019), and an ongoing goal has been to understand how they can be distributed asymmetrically between the two leaflets of the membrane bilayer so that extreme negative membrane curvature can be generated to drive vesicle fission to completion.
We have also been intrigued by another question raised by the elucidated role of these short lipids. Short PC (sPC) is the precursor of short PA and DAG needed for COPI vesicle fission (Park et al., 2019). Thus, as PC synthesis occurs in the ER (Vance and Vance, 2004), how can sPC be delivered from the ER to sites of COPI vesicle formation at the Golgi to support the fission stage? In this study, we find that PITPβ plays a key role in delivering sPC from the ER to sites of COPI vesicle formation at the Golgi to promote the fission stage. By combining cell-based and reconstitution approaches, we achieve a high level of spatial and temporal resolution in explaining how this delivery occurs. Our results advance a new understanding of how PITPβ acts, define a specific circumstance where membrane contact is needed for vesicular transport, and contribute to a general understanding of how intracellular transport carriers are formed.
Results
COPI transport requires membrane contact between the ER and the Golgi
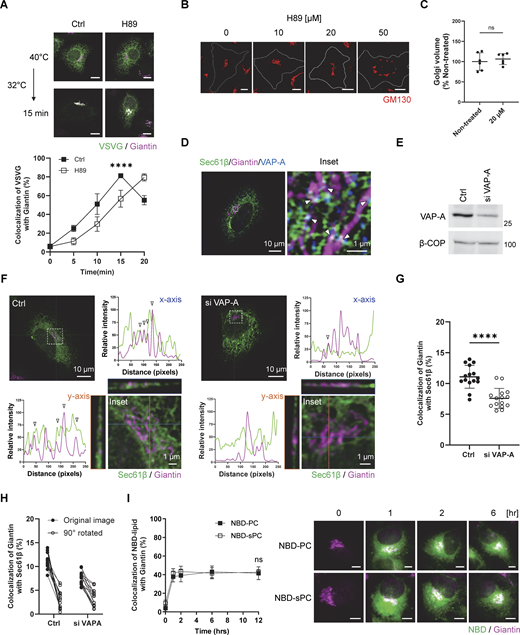
As lipids can be transferred among intracellular membrane compartments by transport carriers or membrane contacts, we initially sought to determine which mechanism is required for sPC delivery that supports COPI vesicle fission. The COPII complex generates carriers from the ER for transport to the Golgi (Brandizzi and Barlowe, 2013; Lee et al., 2004; Suda et al., 2018). As prolonged inhibition of COPII transport disrupts the Golgi complex (Ward et al., 2001), we sought to inhibit COPII transport acutely by treating cells pharmacologically with H89, a compound that inhibits COPII transport while allowing the Golgi to remain intact (Lee and Linstedt, 2000). We first confirmed that this treatment inhibited COPII transport, as tracked by a model secretory protein, vesicular stomatitis virus G protein (VSVG), from the ER to the Golgi (Fig. S1 A). We also confirmed that the dose of H89 treatment allowed the Golgi to remain intact (Fig. S1, B and C). We then found that COPI transport was not inhibited by this treatment (Fig. 1 A), as tracked by the transport of a model COPI cargo, a chimeric protein known as VSVG-KDELR. This cargo accumulates at the Golgi at the permissive temperature (32°C) and becomes transported from the Golgi to the ER upon shifting to the non-permissive temperature (40°C). In this manner, one round of COPI transport can be assessed (Park et al., 2019; Yang et al., 2005, 2008, 2018).
In light of the above results, we next examined whether membrane contact between the ER and the Golgi would be required for COPI transport. The ER forms contacts with other organelles through VAPs expressed on the ER membrane (Murphy and Levine, 2016). As VAP-A has been found previously to participate in forming contact between the ER and the Golgi (Mesmin et al., 2013), we initially confirmed by confocal microscopy that a population of VAP-A colocalized with both an ER marker (Sec61β) and a Golgi marker (giantin) (Fig. S1 D). We next found that siRNA against VAP-A (Fig. S1 E), using a sequence whose specificity had been previously documented (Nthiga et al., 2020), reduced the colocalization of these two markers (Fig. S1, F and G). Furthermore, as specificity control for our colocalization analysis, we rotated one confocal channel by 90°, which also resulted in reduced colocalization, thus confirming that the detected colocalization was not random (Fig. S1 H).
We next found that siRNA against VAP-A inhibited COPI transport (Fig. 1 B). Thus, to gain insight into whether this inhibition could be attributed to impaired sPC delivery, we pursued a functional rescue approach. Lipids can be delivered into cells by incubating them in a culture medium in the presence of albumin (Pinot et al., 2014). We had previously taken this approach to deliver specific lipids to the Golgi (Park et al., 2019). We found that the inhibition of COPI transport induced by siRNA against VAP-A was rescued by feeding cells with PC having shorter (2C10), but not longer (2C18), acyl chains (Fig. 1 C). We also confirmed lipid delivery of both PC lengths to the Golgi by tracking their fluorescence-labeled forms fed to cells (Fig. S1 I). Furthermore, we found that feeding cells with other major lipids of the Golgi in their shorter form (2C10), such as phosphatidylethanolamine (PE) or phosphatidylserine (PS), did not revert the inhibition in COPI transport (Fig. 1 D). Thus, the collective results suggested that sPC delivery needed for COPI transport occurs through membrane contact rather than through COPII transport.
PC transfer by PITPβ promotes COPI transport
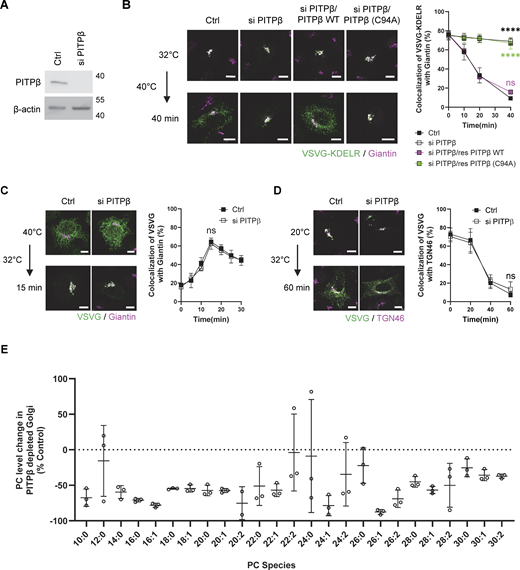
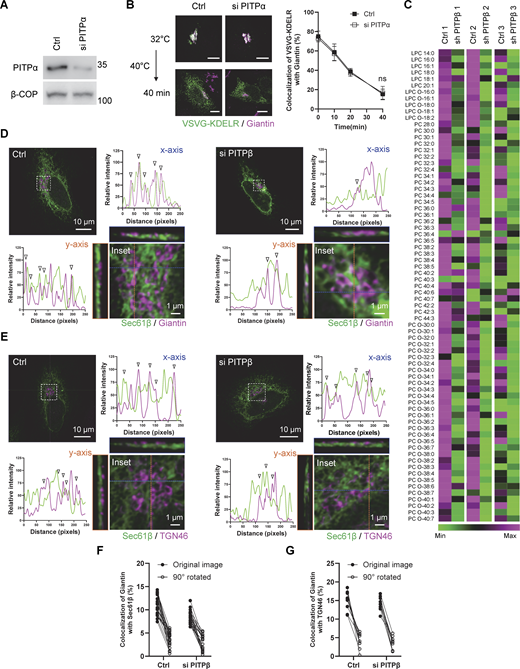
As further confirmation, we next sought to identify a lipid transfer protein predicted to be involved. Phosphatidylinositol transfer protein beta (PITPβ) possesses PC transfer activity (Allen-Baume et al., 2002) and has been found previously to act in COPI transport (Carvou et al., 2010). However, a mechanistic understanding of how it promotes COPI transport has not been achieved. We first confirmed that siRNA against PITPβ (Fig. 2 A) inhibited COPI transport (Fig. 2 B). We also documented the targeting specificity of this inhibition with a rescue experiment (Fig. 2 B). Moreover, the catalytic activity of PITPβ was needed as the catalytic-dead form (C94A) could not rescue inhibition (Fig. 2 B). As controls, we found that this siRNA treatment did not affect anterograde transport into (Fig. 2 C) and out of the Golgi (Fig. 2 D), as tracked through the itinerary of VSVG. We also found that COPI transport was not affected by siRNA against PITPα (Fig. S2, A and B). Thus, inhibition of COPI transport upon targeting against PITPβ showed specificity.
We next collected Golgi membrane from PITPβ-depleted cells to analyze PC levels. Performing lipidomics, we confirmed that the levels of many PC species were reduced (Fig. S2 C). However, this conventional approach of lipidomics, which is untargeted, had difficulty in detecting sPC species (having 28 total carbons or fewer). As sPC species are less abundant than the longer PCs, we next developed a targeted approach, which took into consideration that shorter PCs are less hydrophobic than longer PCs due to the shorter acyl chains. Thus, by isolating the less hydrophobic PCs before performing lipid analysis using mass spectrometry, we were able to detect many sPC species and confirm that many were reduced in Golgi membrane isolated from cells treated with siRNA against PITPβ (Fig. 2 E).
PITPβ also acts in contact formation between the ER and COPI buds at the Golgi
Besides their catalytic activity, lipid transfer proteins have been found to participate in forming membrane contacts (Antonny et al., 2018; Cockcroft and Raghu, 2018; Cohen et al., 2018; Prinz et al., 2020; Reinisch and Prinz, 2021; Venditti et al., 2020; Wong et al., 2019; Wu et al., 2018). Thus, to determine whether PITPβ possesses this additional capability, we initially performed confocal microscopy using Airyscan, which provides twofold greater resolution than the conventional confocal systems. We detected a population of PITPβ colocalizing with both Sec61β (ER marker) and giantin (Golgi marker) (Fig. 3 A). Quantitation revealed a low level of colocalization (Fig. 3 B), which was expected, as colocalization should only occur for regions of the ER and the Golgi that form membrane contact. For this subpopulation, we found that siRNA against PITPβ reduced the colocalization (Fig. 3 B and Fig. S2 D). As a control, we found that siRNA against PITPβ did not reduce membrane contact between the ER and the TGN, as tracked through the colocalization of Sec61β (marking the ER) and TGN46 (marking the TGN) (Fig. 3 C and Fig. S2 E). Further confirming the specificity of these colocalizations, we again rotated one confocal channel by 90° to ascertain that the noted colocalizations were not random (Fig. S2, F and G).
We next sought to detect membrane contact at even higher resolutions. Performing electron tomography, we detected COPI buds to be in closer proximity to ER membranes than the rest of the Golgi membrane (Fig. 3 D). As this finding suggested that the ER could be contacting the Golgi more specifically at sites of COPI vesicle formation, we next sought further support by pursuing proximity ligation analysis. This approach provides greater resolution (detecting proteins within 40 nm of each other) than standard confocal microscopy (detecting proteins up to 200 nm from each other). We first found that siRNA against PITPβ reduced the proximity between calnexin (another ER marker) and giantin (Fig. 3 E). We then tracked COPI buds through ζ-COP and found that siRNA against PITPβ reduced the proximity between calnexin and ζ-COP even more dramatically (Fig. 3 F). As a control, siRNA against PITPβ did not reduce the proximity between calnexin and TGN46 (Fig. 3 G). Thus, these findings suggested that PITPβ acts in contact formation between the ER and the Golgi, with the latter occurring at sites of COPI vesicle formation.
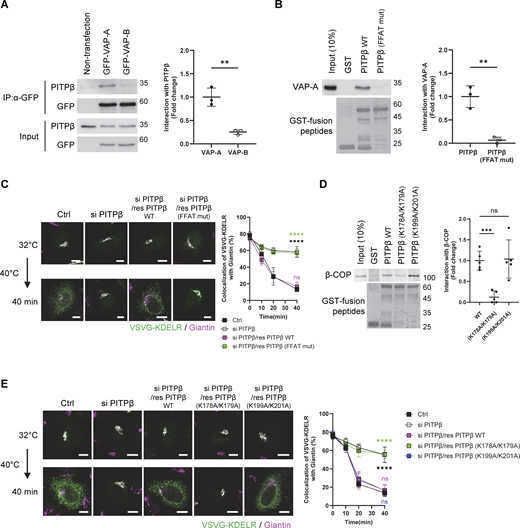
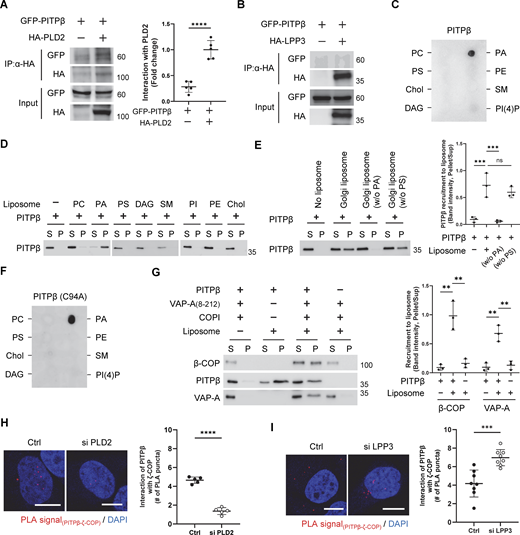
To further characterize how PITPβ acts in this role, we next performed a coprecipitation experiment and found that PITPβ associated with VAP-A in cells, and as control, PITPβ had no appreciable interaction with VAP-B (Fig. 4 A). We then considered that VAPs have been found to interact with binding partners by recognizing an FFAT-like sequence in partner proteins (Murphy and Levine, 2016). As a sequence in PITPβ conforms to this motif, we mutated it (F107A/F108A) and found that VAP-A could no longer interact directly with PITPβ (Fig. 4 B). We also found that replacing the endogenous PITPβ with the mutant PITPβ inhibited COPI transport in cells (Fig. 4 C). This replacement was achieved by performing siRNA against PITPβ followed by rescue using the mutant form. Thus, the results further confirmed a new function for PITPβ, acting to form membrane contact by interacting with VAP-A on the ER membrane.
PITPβ anchors to sites of COPI vesicle formation through multiple interactions
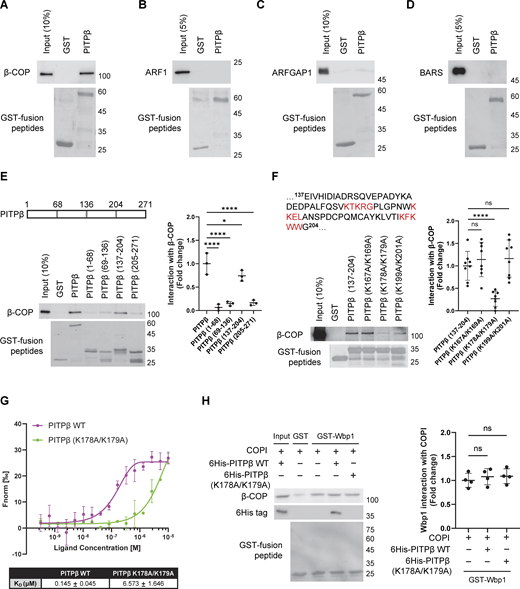
We next sought insight into how PITPβ anchors to the Golgi membrane for this role. Led by our finding above that had revealed the ER to contact the Golgi at sites of COPI vesicle formation, we screened among the COPI-related components by performing pull-down assays using purified proteins. We found that PITPβ could interact directly with coatomer (Fig. S3 A), but not with ARF1 (Fig. S3 B), ARFGAP1 (Fig. S3 C), or BARS (Fig. S3 D). To confirm the functional relevance of the detected interaction, we next sought to identify specific residues in PITPβ needed for binding to coatomer. Initially, we generated truncations and identified a region in PITPβ that interacts directly with the coatomer (Fig. S3 E). As this region contains three di-lysine sequences (Fig. S3 E), and coatomer is known to recognize di-lysine sequences (Cosson and Letourneur, 1994), we next mutated the three candidate sites and identified a single site (K178/K179) in the truncated PITPβ that is critical for its direct interaction with coatomer (Fig. S3 F). We also confirmed that mutating this site prevents the full-length PITPβ from binding directly to the coatomer (Fig. 4 D). Furthermore, we expressed this mutant in cells, which involved siRNA against PITPβ followed by rescue using the mutant form, and confirmed that COPI transport was inhibited (Fig. 4 E).
As the binding of coatomers to di-lysine motifs in cargo proteins has been quantified previously (Ma and Goldberg, 2013), we also sought to quantify coatomer binding to the di-lysine sequence in PITPβ. Pursuing microscale thermophoresis (MST), we found that coatomer bound to wild-type PITPβ (Kd 0.145 µM) better than to the - mutant (Kd 6.573 µM) (Fig. S3 G). Furthermore, we found that coatomer could bind simultaneously to a model COPI cargo (Wbp1) and PITPβ, as assessed by a pull-down assay (Fig. S3 H). Thus, because coatomer has been elucidated previously to bind di-lysine motifs in cargo proteins through its α and β′ subunits (Jackson et al., 2012; Ma and Goldberg, 2013), coatomer is predicted to bind to the di-lysine sequence in PITPβ through a different subunit.
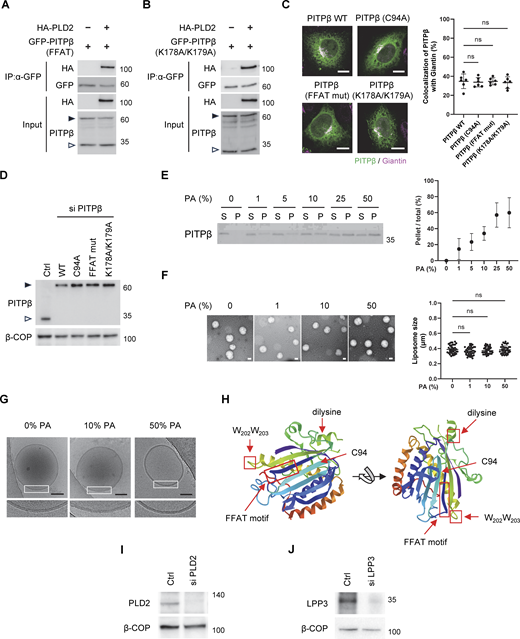
We next pursued another line of investigation that shed further insight into how PITPβ participates in membrane contact. We had found previously that short forms of PA and DAG are needed sequentially for COPI vesicle fission, which involves PLD2 converting sPC to sPA and then LPP3 converting sPA to sDAG (Park et al., 2019). Thus, as PITPβ delivers sPC, we explored the temporal possibility that PITPβ could be acting in membrane contact more specifically during COPI vesicle fission. We initially performed coprecipitation experiments and found that PITPβ interacted with PLD2 in cells (Fig. 5 A). In contrast, PITPβ did not interact with LPP3 (Fig. 5 B). Further, elucidating how PITPβ binds PLD2, we found that the PITPβ mutants (FFAT mutant or K178A/K179A mutant), when transfected and expressed at physiological levels, retained interaction with PLD2 (Fig. S4, A and B). Thus, PLD2 likely binds to a different region of PITPβ than those bound by VAP-A or coatomer. We also confirmed that the PITPβ mutants, when expressed at similar levels, localized to the Golgi membrane similar to the wild-type form (Fig. S4, C and D), thus ruling out that their impaired functions could be attributed to defective membrane localization.
As PLD2 activity generates PA, we next examined whether PITPβ also interacted with PA. As an initial screen, we performed lipid dot-blot analysis and found that PITPβ could bind PA directly (Fig. 5 C). To confirm that this interaction occurred in the context of the membrane, we next generated liposomes and found that PITPβ only bound to liposomes generated using PA (Fig. 5 D). Moreover, as PLD2 has been found to be concentrated at the neck of COPI buds (Park et al., 2019), we found that increasing the level of PA in liposomes led to increased binding by PITPβ (Fig. S4 E). We also confirmed that liposome sizes were similar and the membrane bilayer remained intact in liposomes having higher levels of PA, as assessed by EM (Fig. S4, F and G). We then generated liposomes having a Golgi-like composition of lipids and confirmed that PA was also needed for PITPβ to bind these liposomes (Fig. 5 E). As a control, we found that this binding was not affected when phosphatidylserine, which is another negatively charged phospholipid, was withdrawn from the Golgi-like liposomes (Fig. 5 E). Thus, PITPβ can anchor to the Golgi by binding not only to protein components (coatomer and PLD2) but also to a lipid component (PA) of the COPI bud.
Insight into the portion of PITPβ that binds PA came from a previous study that found a region containing a di-tryptophan (WW) motif to be involved in membrane binding (Shadan et al., 2008). We further noted that this region is distinct from the catalytic domain (Fig. S4 H), which suggests how PITPβ can anchor to the Golgi membrane while also catalyzing PC transfer. Supporting that these two functions involve distinct regions of PITPβ, we found that the catalytic-dead mutant of PITPβ can still bind PA (Fig. 5 F). The structure of PITPβ further suggested that the region responsible for PA binding would be distinct from those involved in binding to VAP-A and the coatomer (Fig. S4 H). Thus, PITPβ could bind simultaneously to PA, coatomer, and VAP-A. As a confirmation, we generated a truncated form of VAP-A that could not bind membrane and found that PITPβ could bind to this form along with coatomer, while also being recruited to PA-containing liposomes (Fig. 5 G).
We also pursued another line of investigation to further confirm that PITPβ binds to the Golgi membrane through PA. COPI vesicle fission involves the sequential activities of PLD2 and LPP3, with the former generating PA and the latter consuming PA (Park et al., 2019). Thus, we examined whether targeting against PLD2 (to reduce PA level at sites of COPI vesicle fission) would reduce the localization of PITPβ at COPI sites, and whether targeting against LPP3 (to increase PA level at sites of vesicle fission) would enhance the localization of PITPβ at these sites. Upon siRNA against PLD2 (Fig. S4 I), we found by the proximity ligation assay that PITPβ showed reduced proximity to ζ-COP (Fig. 5 H). Moreover, upon siRNA against LPP3 (Fig. S4 J), PITPβ showed increased proximity to ζ-COP (Fig. 5 I). These results not only confirmed the importance of PA for PITPβ to bind the Golgi membrane but also further supported that PITPβ forms membrane contact with COPI buds during the fission stage.
Reconstituting lipid transfer and contact formation by PITPβ
As cell-based studies cannot rule out indirect effects, we next sought to reconstitute lipid transfer using purified components to confirm the direct roles of PITPβ. As done previously to reconstitute lipid transfer through membrane contact (Gong et al., 2021; Mesmin et al., 2013), we generated two populations of liposomes, in this case with one having an ER-like composition of lipids and another having a Golgi-like composition. For the ER liposomes, we attached VAP-A onto the membrane using a previously established approach that involves tagging a protein with 6x-histidine for its attachment to liposomes that contain a nickel-bound lipid (Lee et al., 2005). For the Golgi liposomes, we incorporated PA into the liposomal membrane to allow binding by PITPβ. To detect PC transfer from the ER liposomes to the Golgi liposomes, we incorporated NBD-tagged PC and rhodamine-tagged PE into the ER liposomes. When both fluorescence-tagged lipids reside in the ER liposomes, NBD fluorescence is not detectable due to quenching by the rhodamine fluorophore. However, when PC is transferred from the ER liposomes to the Golgi liposomes, dequenching occurs, resulting in the NBD fluorophore becoming detectable (summarized in Fig. 6 A).
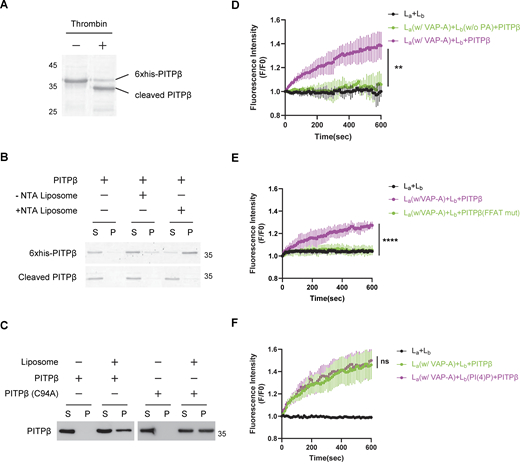
We first generated recombinant PITPβ tagged with 6x-histidine to facilitate its purification. We then removed the 6x-histidine tag from recombinant PITPβ after its purification (Fig. S5 A) and confirmed that the resulting untagged PITPβ could no longer bind liposomes that contained a nickel-bound lipid (Fig. S5 B). Subsequently, when this untagged PITPβ was incubated with ER liposomes and Golgi liposomes, we confirmed contact formation by EM (Fig. 6 B) and also that PC transfer was reconstituted (Fig. 6 C). As a control, transfer did not occur when VAP-A was not attached to the ER liposomes (Fig. 6 C). Transfer also did not occur when a catalytic-dead form of PITPβ was used (Fig. 6 C). In the latter case, we also ruled out that the defect in PC transfer could be attributed to the mutant protein that was unable to bind to the Golgi liposomes, as it bound to these liposomes similar to the wild-type form (Fig. S5 C). PC transfer also did not occur when PA was not incorporated into the Golgi liposomes (Fig. S5 D) or when PITPβ binding to VAP-A was disrupted by using the mutant PITPβ with the FFAT-like sequence mutated (Fig. S5 E).
We next found that PITPβ preferentially transfers PC having shorter acyl chains (Fig. 6 D). Another insight came from the consideration that cholesterol transfer from the ER to the Golgi had been found previously to involve the counter-transfer of PI(4)P (Mesmin et al., 2013). Thus, when also considering that PITPβ transfers not only PC but also phosphatidylinositol (PI) (Allen-Baume et al., 2002), we next incorporated PI into the Golgi liposomes and found that sPC transfer from the ER liposomes to the Golgi liposomes was enhanced (Fig. 6 E). As a control, adding PI(4)P to Golgi liposomes did not affect sPC transfer (Fig. S5 F). We also confirmed more directly the counter-transfer of PI from the Golgi liposomes to the ER liposomes. In this case, we added fluorescence-labeled PI and rhodamine-tagged PE to the Golgi liposomes and observed dequenching upon the addition of PITPβ (Fig. 6 F). Thus, the reconstitution studies not only confirmed the direct roles of PITPβ in catalyzing PC transfer and forming membrane contact but also revealed new findings that PITPβ preferentially transfers shorter forms of PC, and this transfer is enhanced by the counter transfer of PI.
Confirming PITPβ acts in the fission stage through vesicle reconstitution studies
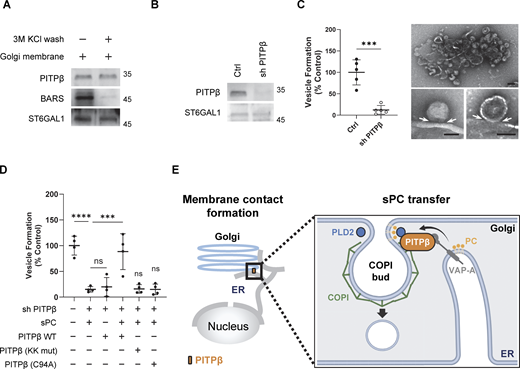
We also pursued the COPI vesicle reconstitution system to confirm more directly that PITPβ is required for COPI vesicle fission. This approach involves the incubation of Golgi membrane with purified factors, as previously described (Park et al., 2015, 2019; Yang et al., 2002, 2005, 2008, 2011, 2018). We initially found that the Golgi membrane contained an appreciable level of PITPβ (Fig. 7 A). As BARS has been shown previously to be released from the Golgi membrane upon high-salt wash (Yang et al., 2005), we tried this approach for PITPβ. However, we did not observe the release of PITPβ (Fig. 7 A). Taking a different approach, we next collected the Golgi membrane that was depleted of PITPβ by stably expressing shRNA against PITPβ in cells and then isolating the Golgi membrane. Immunoblotting confirmed that the isolated Golgi membrane had reduced PITPβ level (Fig. 7 B). We then used this PITPβ-depleted Golgi membrane for the reconstitution system and found that COPI vesicle formation was inhibited (Fig. 7 C). Upon EM examination, we observed the accumulation of buds with constricted necks on the Golgi membrane (Fig. 7 C). Thus, these results confirmed that PITPβ is critical for the fission stage of COPI vesicle formation.
We also confirmed that vesicle reconstitution using PITPβ-depleted Golgi membrane requires sPC to be delivered to the Golgi membrane (Fig. 7 D). For this delivery, we incubated ER liposomes (having sPC and VAP-A on the membrane surface) with PITPβ and Golgi membrane that had been depleted of PITPβ. As controls, vesicle formation was not reconstituted when incubation lacked either VAP-A coating the ER liposomes or PITPβ as a purified factor (Fig. 7 D). Furthermore, when wild-type PITPβ was substituted with mutant forms deficient in binding to coatomer, or catalytic activity, COPI vesicle formation was also not reconstituted (Fig. 7 D). Thus, the results confirmed not only that PITPβ acts specifically at the fission stage of COPI vesicle formation but also its dual role in catalyzing PC transfer and forming membrane contact.
Discussion
We identified a specific circumstance that requires the coordination of membrane contact and vesicular transport. Led by our previous finding that sPC is the precursor of short forms of PA and DAG that are needed for COPI vesicle fission (Park et al., 2019), we initially found that sPC is delivered from the ER to the Golgi through membrane contact rather than through COPII transport. We then found that PITPβ plays a key role in this delivery by not only providing the PC transfer activity but also participating in forming the membrane contact (Fig. 7 E). As PITPβ is not known to act in membrane contact, our findings achieve a new understanding of how PITPβ acts. Furthermore, whereas PITPβ is known to catalyze both PC and PI transfer, we achieve a new understanding of these roles by revealing that both transfers are involved in explaining how PITPβ acts in COPI transport, with PC transfer from the ER to the Golgi enhanced by the counter transfer of PI from the Golgi to the ER.
By combining cell-based studies with reconstitution approaches, we have achieved a high level of spatial and temporal resolution in explaining how sPC is delivered through membrane contact to promote COPI vesicle fission. Spatially, our results reveal an unusual way that a membrane contact can be formed. Rather than two membrane compartments participating in contact formation, a compartmental membrane can also contact a budding carrier for lipid delivery. Temporally, our results suggest that this membrane contact occurs during vesicle fission. Thus, by having sPC being targeted to COPI buds rather than the Golgi membrane more generally, and having this delivery occur at a specific stage of COPI vesicle formation, our findings reveal how these rare lipids can accumulate to relevant levels locally to support COPI vesicle fission.
On the broader level, we note that membrane contact has been found to support vesicular transport, but the precise stage of transport that requires this coordination remains to be better established. Previous studies have implicated that membrane contact between the ER and the Golgi is needed for carrier formation from the TGN (Wakana et al., 2015, 2021). However, evidence supporting this role has been indirect, as only cell-based studies were performed. In contrast, we have pursued the reconstitution of vesicle formation and lipid transfer through membrane contact. Thus, because these approaches provide not only more direct assessments but also higher mechanistic resolution, we achieved a more comprehensive understanding of how membrane contact is needed for a particular stage of vesicular transport. As COPI transport has been a model for elucidating the mechanistic details of vesicle formation, our findings also contribute to the general understanding of how intracellular transport carriers are formed.
Materials and methods
Chemicals and lipids
The following reagents were obtained from GE Healthcare: guanine nucleotide triphosphate (GTP) (10106399001) and protease inhibitor cocktail (P2714) from Sigma-Aldrich, H89 dihydrochloride (sc-3537) and Protein G PLUS-agarose (sc-2002) from Santa Cruz Biotechnology, Ni-NTA agarose from Qiagen (30210), and glutathione Sepharose bead (GE17-0756-01). The following lipids were obtained from Avanti Polar Lipid: dioleoyl phosphatidylcholine (DOPC, 850375), dioleoyl phosphatidylethanolamine (DOPE, 850725), dioleoyl phosphatidylserine (DOPS, 840035), dioleoyl phosphatidic acid (DOPA, 840875), cholesterol (700000), Sphingomyelin (860587), dioleoyl diacylglycerol (DODAG, 800811), dioleoyl phosphatidylinositol (DOPI, 850149), 18:1 phosphatidylinositol-4 phosphate [PI(4)P, 850151], 2C10:0 PC (850325), 2C10:0 PS (840036), 2C10:0 PE (850700), 18:1-12:0 nitrobenzodiazole (NBD)-PC (810133), 14:1-6:0 NBD-PC (810122), 18:1-12:0 NBD-PA (810176), Egg Liss Rhodamine PE (810146), Topfluor-PI (810187), 18:1 DGS-NTA (790404), and 15:0-18:1(d7) PC (791637).
Proteins
Purifications of coatomer, ARF1, ARFGAP1, BARS, and Golgi membrane have been described previously (Park et al., 2016). His tagged PITPβ, BARS, and VAP-A (8-212) in pET-15b were expressed in BL21 cells with isopropyl β-D-1-thiogalactopytanoside (IPTG) induction. Cells were lysed in buffer (20 mM Tris-HCl pH 7.4, 200 mM NaCl, 1% Triton X-100, 1 mM dithiothreitol [DTT], and 10% glycerol) containing protease inhibitor cocktail and then subjected to centrifugation. The resulting supernatant was subjected to purification using a Ni-NTA column. PITPβ and proteins fused to glutathione S-transferase (GST) were subcloned into pGEX-4T3 and then expressed in BL21 cells with IPTG induction. Cells were lysed followed by centrifugation, and then the supernatant was subjected to purification using Glutathione Sepharose beads.
Antibodies
Mouse antibodies against β-COP (M3A5), VSVG (BW8G65), and PLD2, and rabbit antibodies against ARF1, ARFGAP1, and ζ-COP have been previously described (Park et al., 2015, 2019; Yang et al., 2011). The following antibodies were obtained: monoclonal antibody against PITPα and PITPβ (kind gift from Dr. Shamshad Cockcroft, University College London, London, UK); anti-CtBP1/BARS (8684S), anti-HA epitope (3724S), and 6xhis epitope antibodies (9991S) from Cell Signaling; antibodies against GFP (sc-9996), VAP-A (sc-293278), calnexin (sc-46669), and β-actin (sc-47778) from Santa Cruz Biotechnology; antibodies against anti-GM130 (610822) from BD Transduction Laboratories:; mouse anti-LPP3 (ab52581) and rabbit anti-giantin (ab80864) antibodies from Abcam; rabbit anti-TGN46 (PA1-1069) and anti-ST6GAL1 (PA5-106647) antibodies from Invitrogen. Secondary antibodies were obtained from Jackson ImmunoResearch Laboratories, and they include horseradish peroxidase–(HRP) conjugated Goat Anti-Mouse IgG (H+L) (115-035-003), HRP-conjugated Goat Anti-Rabbit IgG (H+L) (111-035-003), HRP-conjugated Goat Anti-Mouse IgG Light chain specific(115-035-174), HRP-conjugated Goat Anti-Rabbit IgG Light chain specific(211-032-171), Cy3-conjugated Goat anti-Mouse IgG (H+L) (115-165-003), Alexa Fluor 488-conjugated Goat Anti-Rabbit IgG (H+L) (111-545-003), and Alexa Fluor 647-conjugated Goat Anti-Rabbit IgG(H+L) (111-605-003).
Plasmids and oligonucleotides
Plasmids encoding for PLD2 in pCGN, HA-tagged LPP3 in pcDNA3.1, GST-tagged Wbp1 tail in pGEX-4T3, VSVG (ts045) in pcDNA3.1, and VSVG(ts045)-KDELR in pROSE have been described (Cole et al., 1998; Cosson and Letourneur, 1994; Gutierrez-Martinez et al., 2013; Yang et al., 2008). PITPβ in pEGFP-C1 was a gift from Dr. Shamshad Cockcroft (University College London, London, UK). PITPβ was subcloned to pET-15b, pcDNA 3.1, and pGEX-4T3 using PCR. Mutant forms of PITPβ were generated using QuickChange II Site-Directed Mutagenesis (Agilent). VAP-A (104447) and VAP-B (104448) in pEGFP-C1 and mEmerald-tagged Sec61β in pEGFP-C1 (90992) were obtained from Addgene. VAP-A(8–212) was subcloned to pET-15b using PCR. The following siRNA sequences were used against targets: PITPα (5′-AACCGTTATTACAAATGAGTA-3′), PITPβ (5′-ACGGAUAUUUACAAACUUCCA-3′), VAP-A (5′-CCUGAGAGAUGAAGGUUUA-3′), PLD2 (5′-GGACAACCAAGAAGAAAUA-3′), LPP3 (5′-GGGACUGUCUCGCGUAUCA-3′) (Carvou et al., 2010; Nthiga et al., 2020; Park et al., 2019). The targeting sequence against PITPβ for shRNA expression (5′-CCGGCCATGTTCTGTTCAGGAGTATCTCGAGATACTCCTGAACAGAACATGGTTTTTTG-3′) was synthesized (VectorBuilder).
Cells
HeLa cells (ATCC), certified as free of mycoplasma contamination, were cultured in Dulbecco’s modified essential medium (Welgene) supplemented with 10% fetal bovine serum (Hyclone), 10 mM HEPES (Gibco), and antibiotics. Transfection of DNA plasmids was performed using GenJet (SignaGen Laboratories). Transfection of siRNA was performed using PepMute (SignaGen Laboratories). 1 µg/ml of DNA plasmids and 30 µM of siRNA were used for transfection. Stable expression of shRNA against PITPβ was established by transduction with lentiviral vector followed by puromycin selection.
In vivo transport assays
Transport assays by tracking the quantitative colocalization of a model cargo with organelle marker in HeLa cells have been described previously (Park et al., 2015; Yang et al., 2018). Briefly, for COPII transport, VSVG(ts045) was transfected and then synchronized at the ER by incubating cells at 40°C for 6 h and then released from the ER by shifting the temperature to 32°C. For COPI transport, VSVG(ts045)-KDELR was transfected and then synchronized at the Golgi by incubating cells at 32°C for 6 h and then released from the Golgi by shifting the temperature to 40°C. For transport from the TGN, VSVG(ts045) was transfected and then synchronized at the TGN at 20°C for 3 h and then released from the TGN by shifting the temperature to 32°C. In each case, cells were fixed at different time points using 4% paraformaldehyde for 10 min and then stained using antibodies against giantin (1:500) and VSVG (1:5) for assays on COPI and COPII transport, and antibodies against TGN46 (1:200) and VSVG (1:5) for transport assay from the TGN. Quantitative colocalizations were then performed (detailed in the next section below). For the functional lipid rescue of COPI transport, a lipid of interest (final concentration, 125 µM) was mixed with DOPC (1:1, molar ratio). The lipid mixture was dried using nitrogen gas and then dissolved in 2 ml of prewarmed culture medium containing bovine serum albumin (BSA) (25 µM). To introduce the lipid to the cells, the lipid-containing medium was incubated with cells for 6 h, followed by the COPI transport assay.
Quantitative colocalization studies
Cells were imaged using a Zeiss LSM900 confocal system with an Airyscan package having a Plan-Apochromat 63× objective and Zeiss URGB (488, 561, and 647 nm) laser lines. Images were imported for processing using the Zen 3.8 software. Images were cropped using Image J and then adjusted for appropriate intensity using Photoshop. The threshold was determined by the Costes method (Costes et al., 2004). Manders coefficient was then obtained, with colocalization expressed as a percentage of one protein population colocalizing with another protein population. For transport assays, five cells were examined individually at each time point per condition per experiment. For colocalization between organelles, eight cells were examined individually per condition per experiment. The number of experiments done is indicated by “n” value specified in the figure legend.
Measuring Golgi volume
HeLa cells were fixed using 4% paraformaldehyde for 10 min and then stained using an antibody against GM130 (1:1,000). For 3D imaging of Golgi, Z-stack images of Golgi were acquired by confocal microscopy with 260-nm Z-step size. Image stacks were concatenated and analyzed using Image J. Golgi volume was then calculated using the 3D Object Counter plugin under the Analyze tab.
Examination of the membrane contacts
Membrane contacts between ER and the Golgi were assessed by two approaches. One approach involves confocal microscopy. Cells were transfected with Emerald-Sec61β using GenJet for 3 days. After fixation using 4% paraformaldehyde, cells were incubated with antibody against giantin to track the cis-Golgi or against TGN46 to track the TGN, followed by quantitative colocalization as described above. Another approach involves the proximity ligation assay (PLA), which uses the DUOlink PLA kit (Sigma-Aldrich) according to the manufacturer’s protocol. Briefly, cells were permeabilized with 0.05% saponin for 20 min and then stained using an antibody that tracks the ER (calnexin, 1:300), the Golgi (giantin, 1:500), the TGN (TGN46, 1:200), or COPI sites at the Golgi (ζ-COP, 1:300) at 4°C for overnight. Cells were then incubated with oligonucleotide-conjugated secondary antibody at 37°C for 1 h, followed by incubation with the ligation solution, which contains ligase and circular oligonucleotide, for 30 min. The signals of polymerized oligonucleotides were then amplified by incubating the cells with an amplification solution containing polymerase at 37°C for 100 min. Cell nuclei were stained using DAPI. PLA signals were detected by confocal microscopy. Image J was used to quantify PLA signals per cell.
Immunoprecipitation and GST pulldown
For immunoprecipitation, cells were lysed with lysis buffer (20 mM Tris-HCl pH 7.4, 200 mM NaCl, 1% Triton X-100 and protease inhibitor). After centrifugation at 17,000 × g for 15 min, the supernatant fraction was incubated with protein G agarose beads that had been bound by the antibody at 4°C for 2 h. Beads were rinsed three times using lysis buffer and subjected to SDS-PAGE followed by western blotting. For in vitro pulldown assays, recombinant GST fusion proteins were expressed in bacteria (BL21 cell) using IPTG induction. Cells were lysed using lysis buffer and centrifuged at 4,000 × g for 15 min. The supernatant fraction was then incubated with glutathione Sepharose beads (GE healthcare). GST fusion proteins on beads were incubated with soluble recombinant proteins at 25°C for 2 h in reaction buffer (25 mM Tris-HCl pH 7.4, 50 mM KCl, 2.5 mM Mg(OAc)2, 1 mM DTT, and 0.1% Triton X-100). Beads were washed twice using reaction buffer and then subjected to SDS-PAGE followed by western blotting.
Lipid dot blot
Purified lipids (200 nmol) were dissolved in chloroform:methanol:50 mM hydrochloric acid (HCl):ponceau S (250:500:200:2, vol/vol/vol/vol), spotted on the nitrocellulose membrane (10600002; GE Healthcare), and then dried for 1 h in a dark room. After blocking with 3% BSA in TBS (30 mM Tris-HCl pH7.4, 100 mM NaCl) for 2 h, the membrane was incubated with recombinant PITPβ (1 µg/ml) overnight at 4°C. The membrane was washed three times using TBST buffer (30 mM Tris-HCl pH7.4, 100 mM NaCl, 0.05% Tween-20) and then probed by immunoblotting.
Liposome binding assay
Liposomes were generated using a mixture of pure lipids. To mimic the Golgi composition, the mixture used was DOPC (50%), DOPE (10%), cholesterol (17%), sphingomyelin (8%), DOPS (7%), DOPI (6%), DOPA (1%), and DODAG (1%). To mimic the ER composition, the lipid mixture was DOPC (70%), DOPE (25%), and DOPS (5%). For simplified liposomes, the mixture was lipids of interest (30%) and DOPC (70%). Lipid mixture (200 µg) was evaporated using N2 gas and then resuspended in 200 µl of reaction buffer (25 mM Tris-HCl pH 7.4, 50 mM KCl, 2.5 mM Mg(OAc)2) for hydration at room temperature (RT) overnight and then passed through a 400-nm filter membrane in a mini-extruder (Avanti Polar Lipids). Liposome binding studies were performed by incubating liposomes (20 µg) with 1 µM of soluble recombinant protein of interest in 100 µl of reaction buffer (25 mM Tris-HCl pH 7.4, 50 mM KCl, 2.5 mM Mg(OAc)2) at room temperature for 30 min, followed by centrifugation at 200,000 × g at 4°C for 30 min. The pellet and supernatant fractions were then assessed by SDS-PAGE followed by western blotting to assess the membrane-bound versus the soluble fraction, respectively.
EM imaging of PA-containing liposomes
Liposomes were generated using a mixture of pure DOPC lipids with various concentrations of DOPA lipids (100% DOPC, 99%DOPC/1%DOPA, 90%DOPC/10%DOPA, or 50%DOPC/50%DOPA). Lipid mixture (100 µg) was then evaporated using N2 gas and resuspended in 100 µl of reaction buffer (25 mM Tris-HCl pH 7.4, 50 mM KCl, 2.5 mM Mg(OAc)2) for hydration at room temperature (RT) overnight and then passed through a 400-nm filter membrane in a mini-extruder (Avanti Polar Lipids). For TEM sample preparation, the liposome solution was placed onto a carbon grid (CF200-CU; Electron Microscopy Science). The grid was rinsed twice with distilled water and then stained using 1% uranyl acetate, followed by examination using a JEOL JEM-1011 electron microscope. For cryo-EM imaging, 3.2 μl of liposome solution was applied to a glow-discharged Quantifoil 1.2/1.3 copper EM grid. After a 30-s incubation, the grids were blotted for 5 s at 18°C and 100% humidity using a Vitrobot Mark IV (Thermo Fisher Scientific) and then immediately plunge-frozen in liquid ethane. Cryo-EM images were collected on a Titan Krios G4 (Thermo Fisher Scientific) operating at 300 kV, with images recorded at 130,000× magnification and a total dose of 50 e-/Å2.
Electron tomography
A cell slurry (cells resuspended in 20% BSA/PBS) was transferred to B-type planchettes (3 mm) and rapidly frozen with a Leica EM ICE high-pressure freezer (Leica Microsystems). The frozen specimens were freeze-substituted in anhydrous acetone containing 2% osmium tetroxide (OsO4) at −80°C for 72 h in an AFS2 machine (Leica Microsystems). After being slowly warmed to room temperature for over 48 h, samples were rinsed with 100% acetone three times to remove excess OsO4. Cell pellets were separated from planchettes and gradually infiltrated into Embed-812 resin (14120; Electron Microscopy Sciences) in a dilution gradient (10%, 25%, 50%, 75%, 100% in acetone) over 2 days. The samples in 100% resin were polymerized at 60°C overnight. For electron tomography analysis, serial-section ribbons were collected on copper slot grids (150–200 nm) and post-stained. The ribbons were sandwiched between two layers of formvar film and coated with 15 nm gold particles (Ted Pella). Tilt series from ±60 at 1.5° intervals were collected in the brightfield transmission electron microscopy (TEM) mode with a 200-kV Tecnai F20 intermediate voltage electron microscope (Thermo Fisher Scientific). Tilt series around two orthogonal axes were acquired from each section. The IMOD (University of Colorado Boulder) software package was used for tomographic reconstruction as described previously (Mai and Kang, 2017; Toyooka and Kang, 2014). The distance between the endoplasmic reticulum (ER) and the Golgi membranes was measured using the IMOD software package. Open contours consisting of two points (i.e., straight lines) were drawn to span the shortest distances between the ER and Golgi membranes. The lengths of the line models in the tomograms were extracted with the imodinfo command to calculate their average and standard deviation values.
Reconstitution of COPI vesicle formation
Reconstitution of COPI vesicle formation was performed as previously described (Park et al., 2016) with minor modification. Briefly, salt-washed (3 M KCl) Golgi membrane (100 µg) was pelleted by centrifugation at 17,000 × g at 4°C for 30 min. The pellet containing Golgi membrane was resuspended using 1 ml of washing buffer (25 mM HEPES-KOH pH 7.4, 3 M KCl, 2.5 mM Mg(OAc)2, 1 mg/ml soybean trypsin inhibitor), followed by incubation on ice/water for 5 min and then incubated with myristoylated ARF1 (6 µg/ml), ARFGAP1 (6 µg/ml), BARS (3 µg/ml), coatomer (6 µg/ml), and GTP (2 mM) at 37°C for 30 min in 300 µl of reaction buffer (25 mM HEPES-KOH pH 7.4, 50 mM KCl, 2.5 mM Mg(OAc)2, 1 mg/ml soybean trypsin inhibitor, 1 mg/ml BSA, and 200 mM sucrose). After incubation, centrifugation was performed at 17,000 × g at 4°C for 30 min to pellet Golgi membrane. Centrifugation was then performed on the supernatant at 200,000 × g using TLA120.2 rotor at 4°C for 30 min to pellet vesicles. The level of reconstituted COPI vesicles was determined by western blotting using the antibody against β-COP. For PITPβ rescue experiments, recombinant PITPβ (18 µg/ml) was added along with the purified protein factors during the incubation.
Reconstitution of lipid transfer and contact formation
The reconstitution system was performed similar to that described previously (Gong et al., 2021; Mesmin et al., 2013), with slight modifications. In brief, ER-like (donor) liposomes (La: 64% DOPC, 21% DOPE, 6% DGS-NTA, 5% DOPS, 2% rhodamine and 2% NBD-labeled PC with different chain length; La (PI): 44% DOPC, 21% DOPE, 20% DOPI, 6% DGS-NTA, 5% DOPS, 2% rhodamine and 2% NBD-PC), and Golgi-like (acceptor) liposomes (Lb: 53% DOPC, 18% cholesterol, 11% DOPE, 8% sphingomyelin, 5% DOPS, and 5% DOPA; Lb (PI): 33% DOPC, 20% DOPI, 18% cholesterol, 11% DOPE, 8% sphingomyelin, 5% DOPS and 5% DOPA; Lb PI(4)P: 33% DOPC, 20% PI(4)P, 18% cholesterol, 11% DOPE, 8% sphingomyelin, 5% DOPS and 5% DOPA) were prepared. To track PI transport from Golgi liposomes to ER liposomes, Golgi-like (donor) liposome (Lb: 51% DOPC, 18% cholesterol, 9% DOPE, 8% sphingomyelin, 5% DOPS, 5% DOPA, 2% Rhodamine-PE and 2% Topfluor-PI) and ER-like (acceptor) liposome (La: 66% DOPC, 23% DOPE, 6% DGS-NTA and 5% DOPS) was prepared. ER-like liposomes (20 µg) were then incubated with 2 µM of 6x his tagged VAP-A (aa 8-212) at room temperature for 30 min. For lipid transfer and contact formation, donor liposomes (10 µg) were incubated with acceptor liposomes (3 µg) at 37°C in the presence of recombinant PITPβ (1 µM). Lipid transfer from donor liposome to acceptor liposome was measured by fluorescence signal of fluorophore-tagged lipid using Synergy H1 microplate reader (Excitation: 463 nm and Emission: 536 nm).
EM examination of liposomes in the lipid transfer assay
The ER-like liposome (La: 66% DOPC, 21% DOPE, 6% DGS-NTA, 5% DOPS, and 2% rhodamine-PE), Golgi-like liposome (Lb: 51% DOPC, 18% cholesterol, 11% DOPE, 8% sphingomyelin, 5% DOPS, 5% DOPA, and 2% NBD-PA) were prepared. ER-like liposomes (10 µg) were then incubated with 2 µM of 6x his tagged VAP-A (aa 8–212) at room temperature for 30 min. For contact formation, VAP-A–attached ER-like liposomes were then incubated with Golgi-like liposomes (10 µg) in the presence of recombinant PITPβ (1 µM) for 5 min at RT. Liposomes were then imaged using a JEOL JEM-1011 electron microscope.
Lipid analysis of Golgi membrane by conventional (untargeted) lipidomics
Golgi membrane was extracted using a modified Folch method as previously described (Matyash et al., 2008). Briefly, 200 µl of methanol and 600 µl of methyl tert-butyl ether (MTBE) containing SPLASH lipidomix internal standard mixture (Avanti Polar Lipids) were added into 350 µg of Golgi sample. After mixing for 1 h, 200 µl water was added for phase separation followed by centrifugation at 13,000 rpm for 10 min. The upper organic phase, MTBE solution containing the lipid extracts, was collected and dried with nitrogen gas. The dried lipid extracts were stored at 4°C and then resuspended in 150 µl of isopropyl alcohol/acetonitrile/water (65:30:5 vol/vol/vol) for UPLC-MSE analysis, which used the same amount of internal standard lipid such as 1-pentadecanoyl-2-oleoyl(d7)-sn-glycero-3-phosphocholine (15:0-18:1(d7) PC) in SPLASH Lipidomix internal standard mixture. The extracted lipids were analyzed using a Waters Synapt G2 HDMS mass spectrometer (Waters), which was operated on the MassLynx 4.1 software and coupled to the AQUITY UPLC system. The chromatographic separation was performed on a CSH C18 AQUITY UPLC column (1.7 μm, 2.1 mm × 100 mm; Waters) with the mobile phases A and B, which consisted of acetonitrile/water (60:40, vol/vol) and 2-propanol/acetonitrile (90:10, vol/vol), with 0.1% formic acid and 10 mM ammonium formate. The buffer for mobile phase B was mixed for 3 h to fully dissolve. The flow rate and injection volume were 0.4 ml/min and 5 µl, respectively. The gradient of mobile phase B was as follows: 0 min, 40%; 2 min, 43%; 2.1 min, 50%; 12 min 54%; 12.1 min, 70%; 18 min, 98%; 18.1 min, 40% and 20 min, 40%. The total chromatographic separation run-time was 20 min. We prepared a pooled QC sample at the beginning of the study, and aliquots of the same pooled QC sample were used for the entire study. QC samples were measured first and last, and for every six samples. The all ion fragmentation spectra were obtained in positive-ion modes of the ESI source. The MS conditions were optimized as follows; i.e., the capillary voltages were 3 and 1 kV, the cone voltage was 40 V, the source temperatures were 140°C, and the acquisition mass range of m/z was 50–1,200 for positive ionization modes. The high-energy MS spectra were acquired alternatively with the low-energy MSE spectra in UPLC/MSE analysis. The collision energy used for acquiring the high-energy and low-energy MS spectra was 35 and 6 eV, respectively. The high-energy MS spectra provided information on lipid structures. Lipids were annotated using MS-DIAL version 5.1.230429 (Tsugawa et al., 2015). The relative value of each lipid was calculated and visualized as a heatmap using Morpheus software (Broad institute).
Detection of sPCs by targeted lipidomics
Golgi membrane (6 mg) was dissolved in 3 ml of methanol:chloroform (2:1, vol/vol) containing SPLASH Lipidomix internal standard mixture (Avanti Polar Lipids). After adding 1 ml of chloroform and 1.3 ml of 0.1 M of HCl, the mixture was centrifuged at 1,500 × g for 10 min. The lower phase containing lipids was collected and evaporated using nitrogen gas and then reconstituted in methanol chloroform solution (9:1, vol/vol). PC species were detected by UHPLC coupled electrospray ionization tandem mass spectrometry using an AB Sciex 6,500+ Q-trap mass spectrometer system operated in negative ionization/multiple reaction monitoring mode. Lipids were separated by hydrophilic-interaction chromatography using a Luna 3um NH2, 2 × 100 mm column (Phenomenex Inc) and detected using lipid species–specific precursor/product ion pairs. The column was equilibrated using solvent A (93% acetonitrile, 7% dichloromethane, and 2 mM ammonium acetate). Samples were eluted using a gradient with solvent A and solvent B (50% acetonitrile, 50% water, 2 mM ammonium acetate pH 8.2) as follows: 0–0.01 min: 0–10% of solvent B, 0.01–9 min: 10–50% of solvent B, 9–10.1 min: 50–100% of solvent B, and 10.1–12.5 min: 100% of solvent B, at a flow rate of 0.6 ml/min. PC species were detected in negative ionization mode as their acetate adducts. Peaks were identified, integrated, and reported using AB Sciex OS-Multiquant software. Integrated peak areas for the individual PC species were normalized to the phosphatidylcholine internal standard.
Microscale thermophoresis analysis
The interaction between recombinant PITPβ and coatomer was quantified by microscale thermophoresis (MST) using a Monolith NT.115 pico instrument (NanoTemper Technologies). PITPβ was fluorescently labeled using Alexa Fluor 647 NHS ester (Succinimidyl ester) (Invitrogen) for 1 h at room temperature in 100 mM NaHCO3 pH 8.3. The degree of labeling was determined by measuring the fluorophore absorption at 647 nm. A 16-point serial dilution series of the unlabeled coatomer was prepared in TBS (20 mM Tris-HCl pH 7.4, 150 mM NaCl) spanning concentrations from 0.3 nM to 10 µM. The labeled PITPβ was kept at a constant concentration of 10 nM. After a 5-min incubation at room temperature, the samples were loaded into Monolith NT.115 capillaries (NanoTemper Technologies). Measurements were performed using an MST power determined by autodetect mode at 25°C. The normalized fluorescence (Fnorm) was calculated from the ratio of fluorescence signals between heated and non-heated states. The binding curve was generated using GraphPad Prism version 10.3.1 (GraphPad Software).
Statistical analysis
Sample sizes are indicated in the figure legends. The sample size used was based on our previous familiarity with the assays. For comparison between the two conditions, significance was tested by the paired two-tailed Student’s t test, using Excel or Prism software.
Online supplemental material
There are five supplemental figures. Fig. S1 further characterizes the roles of COPII transport and VAP-A–mediated contact formation in sPC transfer. Fig. S2 further characterizes the role of PITPβ in COPI transport and contact formation. Fig. S3 further characterizes how PITPβ binds to coatomer and VAP-A. Fig. S4 further characterizes how PITPβ binds to other key factors involved in COPI transport. Fig. S5 further characterizes lipid transfer and contact formation by PITPβ.
Data availability
Primary data are available from the corresponding authors upon reasonable request.
Acknowledgments
We thank K. Mai for electron microscopy technical advice and S. Cockcroft for advice and reagents related to PITPβ.
This work was funded by grants to S.-Y. Park (National Research Foundation of Korea: RC-2020-NR050842, RS-2023-00208127, and RS-2024-0344154, Suh Kyungbae Foundation: SUHF-24010035, Samsung Science and Technology Foundation: SRFC-MA2402-13), to V.W. Hsu (National Institutes of Health: R37GM058615), to A.J. Morris (Department of Veterans Affairs, IK6 BX006469), and to B.-H. Kang (Hong Kong Research Grant Council: GRF14113921).
Author contributions: K. Park: Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, S. Ju: Formal analysis, Investigation, H. Choi: Investigation, Writing - review & editing, P. Gao: Formal analysis, Investigation, Methodology, G. Bang: Investigation, J.H. Choi: Investigation, J. Jang: Project administration, Supervision, A.J. Morris: Conceptualization, Investigation, Methodology, Resources, Supervision, Visualization, Writing - review & editing, B.-H. Kang: Investigation, Methodology, Supervision, Visualization, V.W. Hsu: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing - original draft, Writing - review & editing, S.-Y. Park: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Visualization, Writing - original draft, Writing - review & editing.