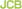The yeast vacuole (similar to the lysosome in higher eukaryotes) is the recipient of cargo from both the secretory and endocytic pathways. Studies of this highly dynamic organelle have been crucial to our general understanding of intracellular transport, and Christian Ungermann continues to elucidate the many different steps of vacuole fusion and the proteins that regulate them.
After studying biochemistry at the University of Tübingen, Ungermann earned his PhD with Walter Neupert and discovered that the molecular chaperone Hsp-70 drives mitochondrial protein import (1). As a postdoc in Bill Wickner's laboratory at Dartmouth Medical School, he switched his attention to the vacuole, helping to define the key sequential steps of membrane fusion: priming, tethering, and docking (2, 3). He's continued on this path as a group leader, first in Heidelberg and now as a full professor at the University of Osnabrück, recently discovering a new tethering complex called CORVET that, along with the HOPS complex, regulates traffic between the vacuole and endosomes (4). These multi-subunit complexes form the initial contact between membranes, acting as downstream effectors of small GTPases called rabs. In addition, his laboratory is interested in the regulation of vacuole fusion by post-translational modifications, including phosphorylation and palmitoylation (5, 6).

In a recent interview, Ungermann spoke about his career, the power of yeast in understanding cell biology, and which steps of membrane fusion will be dissected next.
LOOKING FOR THE ANSWERS
When did your interest in science begin?
I think it started toward the end of high school, when I was generally interested in biology, but wasn't satisfied with the simple answers I got. I decided to move more into the field of biochemistry, and there it started to become really interesting. So initially I was like lots of children who were just interested in biology as a phenomenon.
At which point in your studies did you realize that research was for you?
I was always blessed with being quick at doing experiments, and I really enjoyed having hands-on experience. But it was really when I went to the US, for a master's degree at Oregon State University. I went over with an exchange program and was then exposed to the rather research-oriented American system. We were confronted with lots of papers that we had to read and assignments to write, and we had to explore the literature in a lot more detail. That was far beyond what I did before.
What made you choose Walter Neupert's laboratory for your PhD?
When I was looking for PhD positions, my girlfriend—now my wife—and I decided that we would like to go back to Germany after being in the US for two years. I was always fascinated about mitochondria and mitochondrial import.
This was before the Internet was really present, so you had to go to the library, look at papers, look at records, and try to figure out whether this is something for you.
That must have been a lot of work compared with how it is nowadays.
It's kind of difficult to describe to people who haven't had this experience! One of the things I was completely excited about was that the Neupert laboratory could reconstitute a system of transportation from purified organelles and purified components. I found this very appealing.
SWITCHING CONTINENTS, TOPICS
Were you tempted to stay with mitochondrial import for your postdoc?
Yes, I thought about it. When you leave the laboratory you always think everything is explored, and then you look at the mitochondrial literature ten years later, and they found out so many more components and pathways! You can never predict how a field evolves.
But I always switch a little bit in my interests. I became more and more fascinated with vesicle fusion at that time, through the talks I listened to during my PhD. So even though I found mitochondrial import very exciting, I felt that this was actually the direction that would be fruitful for me. The vesicular transport field was really exploding. Randy Schekman and Jim Rothman had already explored the basic biochemistry and the genes involved.
This is around the time when SNARE proteins were first suggested to control membrane fusion?
Yes, that came up in 1993, and was further looked at in ’94, and there was a sort of tapering off period where people didn't really progress much beyond the initial observations. Then the vacuole fusion field showed up and suddenly there was a system that was accessible where you could target the components again. With mitochondria you had purified organelles that you could work with and manipulate, and I felt that vacuoles were a similarly attractive system.
Did you think of staying in the US after your postdoc?
Yes. It looks like I'm switching between continents continuously! Bill Wickner offered to support me in applying to the American academic communities. But, for me I felt like I was more European, that this was my cultural home. I thought about it for a little while, but we had three children—twin girls and a son—and so we felt that, for us, the better choice was to go back to Germany.
How would you compare the academic systems in the US and Germany?
The German system doesn't have a tenure track, but it's slowly evolving. Instead Germany has a habilitation system, in which basically you have to summarize the research that you've done up to a certain point, and you have to defend it against an audience of scientists. Also, you have to show that you have done enough teaching. In most cases we have to reapply for a position, and it's rarely the case that it's at the same university. Most of the time you have to move. So I guess that's the main difference from the US system. In the US, you have a very low teaching obligation initially, and you have the tenure system.
How do you manage to balance your teaching and research responsibilities?
It's always a matter of juggling. You try to do it as best as you can, and it works out, I have to say. It's also an enjoyable part of my position: to interact with young students and talk to them about science, and try to give them some ideas. I feel that this is actually also training me because I have to go back and explore the things that I do in the laboratory and see how they compare to what I teach.
And you're a father, so how do you balance family and research?
There's a certain limitation of time, but we do lots of things as a family, like rock climbing, skiing, and hiking. We go to the theater and movies, like everyone else. My children were born in the first year of my PhD. I think it's possible to balance the two. There's nothing that excludes it. But I had the advantage that my wife took a good part of the child-raising work on her shoulders. Otherwise, I wouldn't have had the chance to work at this level, and at this intensity.
THE POWER OF YEAST
As a model organism, are yeast as useful now as they ever have been?
I think people tend to question this to some degree now, but if you consider that more than 90% of genes are the same between human and yeast, it's basically a question of which system is best to address your specific scientific question. Working with the HOPS and CORVET complexes, these genes are conserved, and we have much better access to addressing their specific function in yeast, since we can purify the different complexes.
We also have the advantage of not having so many isoforms that can compensate for each other. The complexity is greater if you go to a higher eukaryote.
So it's the combination of good genetics and good biochemistry?
I suppose that is the advantage. It's always pleasing and surprising at the same time to see that a lot of fundamental processes—be it the endocytic pathway or the cell cycle—are conserved across eukaryotes. For the questions that we address, we feel that yeast is ideal, since it provides us with a very nice assay system, which in mammalian cells actually becomes more tricky.
What are you working on at the moment?

The endosomal protein Vps21 (green), is concentrated close to the vacuole (red) when a tethering protein is overexpressed.
The endosomal protein Vps21 (green), is concentrated close to the vacuole (red) when a tethering protein is overexpressed.
We're trying to dissect the CORVET complex and really get an idea of how the initial steps of membrane fusion are working. We believe that the CORVET, like the HOPS complex, combines a few activities. One activity is certainly effector function for the endosomal rab GTPase, Vps21. In addition, it should include a nucleotide exchange function to activate the rab to begin with. And we would expect that it has an ability to bind a SNARE as well, so that it can regulate downstream fusion events. We're trying to dissect these different steps.
We're also trying to look more at the enzymatic mechanism of protein palmitoylation as a regulatory step in fusion. It's actually surprising that the substrate specificity of the palmitoylation machinery is rather low—it can actually modify a lot of proteins. We also don't know how the palmitate is lost. People have suggested it might be done by thioesterases, but at least the yeast protein can be deleted without much of an effect. Maybe the mechanism is a lot different to what we've been thinking.