
1-20 of 120308
Follow your search
Access your saved searches in your account
Would you like to receive an alert when new items match your search?
Journal Articles
Journal:
Journal of Cell Biology
J Cell Biol (2026) 225 (9): e202601131.
Published: 07 August 2026
Includes: Supplementary data

in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
Figure 1. Altered transcriptome in STAT5A KO HeLa cells. (A) PCA of RNA-seq expression profiles. Two-dimensional PCA plot displaying the variance across samples based on normalized gene expression. Points represent individual samples. Colors More about this image found in Altered transcriptome in STAT5A KO HeLa cells. (A) PCA of RNA-seq expressi...
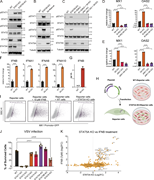
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
Figure 2. STAT5A KO leads to constitutive activation of the IFNAR-JAK-STAT pathway via autocrine type I IFN production. (A–C) Western blot analysis of STAT levels without external type I IFN addition. (A) Analysis of STAT1, STAT2, STAT3, More about this image found in STAT5A KO leads to constitutive activation of the IFNAR-JAK-STAT pathway vi...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
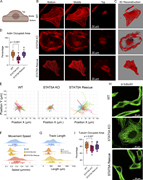
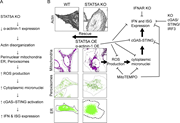
Figure 3. STAT5A KO leads to reorganization of the actin network. (A–C) Actin networks observed by confocal microscopy. (A) Schematic representation of the optical slice positions. (B) Confocal images of WT, STAT5A KO, and STAT5A rescue More about this image found in STAT5A KO leads to reorganization of the actin network. (A–C) Actin networ...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
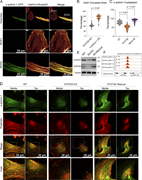
Figure 4. α-Actinin-1 overexpression restores actin bundle formation in STAT5A-deficient cells. (A) Confocal microscopy images showing STAT5A KO cells overexpressing LifeAct-mScarlet3 and α-actinin-1–GFP. Box 1 and Box 2 highlight cells lacking More about this image found in α-Actinin-1 overexpression restores actin bundle formation in STAT5A-defici...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
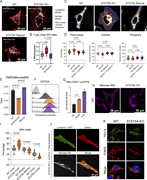
Figure 5. Disrupted actin cytoskeleton in STAT5A KO cells impacts mitochondrial organization. (A) Confocal microscopy images of WT, STAT5A KO, and STAT5A rescue cells overexpressing LifeAct-mScarlet3 and stained with MitoView 650. (B) Boxplot More about this image found in Disrupted actin cytoskeleton in STAT5A KO cells impacts mitochondrial organ...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
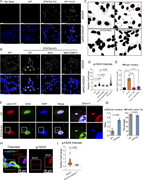
Figure 6. STAT5A KO results in genomic DNA damage. (A) WT and STAT5A KO cells were either untreated or treated with 100 µM H2O2 for 1 h. Cells were stained with anti-γH2AX to detect DNA damage. (B) WT and STAT5A KO cells, either untreated or More about this image found in STAT5A KO results in genomic DNA damage. (A) WT and STAT5A KO cells were e...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
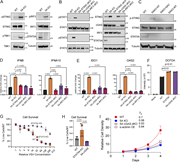
Figure 7. cGAS-STING pathway is activated by cytosolic DNA in STAT5A KO cells. (A–C) Western blot analysis of cGAS-STING pathway activation. (A) Phosphorylated STING, TBK1, and IRF3 in WT and STAT5A KO cells. (B) pSTAT1 and pSTAT2 levels in More about this image found in cGAS-STING pathway is activated by cytosolic DNA in STAT5A KO cells. (A–C) ...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
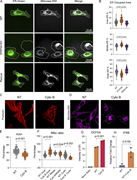
Figure 8. Perinuclear distribution of cellular organelles is a general phenomenon of actin disruption. (A) Representative confocal fluorescence microscopy images showing the spatial distribution of the ER and mitochondria in WT, STAT5A KO, and More about this image found in Perinuclear distribution of cellular organelles is a general phenomenon of ...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
Figure 9. STAT5A KO B16F1 cells recapitulate the HeLa phenotype. (A) Flow cytometry quantification of ROS in unstained control (None), in comparison with DCFDA-stained WT B16F1 cells and B16F1 STAT5A KO cells. (B) RT-qPCR analysis of mIFN-β More about this image found in STAT5A KO B16F1 cells recapitulate the HeLa phenotype. (A) Flow cytometry ...
in STAT5A–α-actinin axis links actin integrity to mitochondrial homeostasis and innate immune activation
> Journal of Cell Biology
Published: 07 August 2026
Figure 10. Schematic model illustrating the downstream consequences of STAT5A KO. (A) Simplified flow diagram summarizing the cascade of events following STAT5A loss. (B) Experimental framework supporting each step of the model. STAT5A KO More about this image found in Schematic model illustrating the downstream consequences of STAT5A KO. (A) ...
Journal Articles
Journal:
Journal of Cell Biology
J Cell Biol (2026) 225 (10): e202510098.
Published: 06 August 2026
Includes: Supplementary data
in Characterizing the membrane recruitment domain of BDCPs and its role in gray platelet syndrome
> Journal of Cell Biology
Published: 06 August 2026
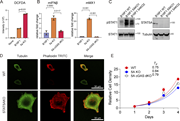
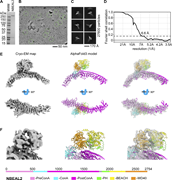
Figure 1. Cryo-EM analysis of NBEAL2. (A) SDS-PAGE gel of purified NBEAL2 protein. (B) Representative cryo-EM micrograph of the NBEAL2 preparation with particles contributing to 2D classes in green circles. Scale bar is 50 nm. (C) 2D class More about this image found in Cryo-EM analysis of NBEAL2. (A) SDS-PAGE gel of purified NBEAL2 protein. ...
in Characterizing the membrane recruitment domain of BDCPs and its role in gray platelet syndrome
> Journal of Cell Biology
Published: 06 August 2026
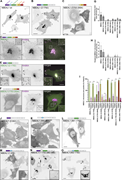
Figure 2. Mapping the membrane-recruitment domain of NBEAL1. (A–C) Representative confocal images of HeLa cells stably expressing 3xFlag-EGFP-tagged full-length or indicated deletion mutants of human NBEAL1. NBEAL1-positive Golgi are marked by More about this image found in Mapping the membrane-recruitment domain of NBEAL1. (A–C) Representative co...

